The Atacama Desert: A World Apart
The Harsh Origins of Copiapoa
To appreciate the singular beauty of Copiapoa, first consider the environment that shaped them: Chile’s Atacama Desert.
Covering 40,500 square miles, the Atacama is the oldest and driest non-polar desert on Earth. It experiences some of the highest measured surface solar irradiance on the planet, and parts have gone centuries without recorded rain. Despite the extremity, the coastal Atacama is classed as a cool desert, a paradox that defines its uniqueness.
Geography and extreme climate
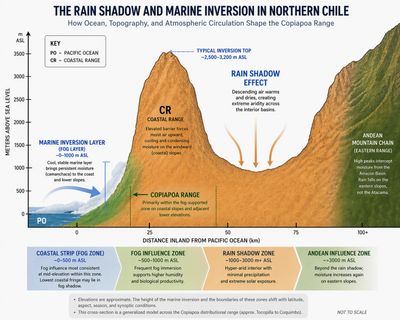
Stretching along Chile's northern Pacific coast, the Atacama Desert begins at the ocean itself. The land rises abruptly from the Pacific as the steep Coastal Range, then drops inland into the desert interior before rising again toward the Andes. This compressed topography creates one of the sharpest environmental gradients on Earth.
Persistent high-pressure over the Pacific suppresses rainfall, while the cold Humboldt Current stabilizes the lower atmosphere, producing frequent coastal fog but little precipitation. Inland, the Coastal Range blocks moisture penetration, and the Andes impose a powerful rain shadow from the east. Together, these forces generate extreme hyper-aridity while maintaining a narrow, fog-fed coastal margin.
So otherworldly is the Atacama Desert that NASA tests instruments here, and the skies host world-class observatories. For Copiapoa, this is not a hostile void but home: adaptation to intense light, mineral soils, and extreme dryness makes them inseparable from this desert world.

The Atacama Desert in Northern Chile
Extreme Solar Irradiance in the Atacama
The Atacama Desert experiences some of the highest levels of surface ultraviolet radiation measured anywhere on Earth. High elevation, persistent cloudless skies, and a relatively low ozone column combine to create an exceptionally intense solar environment. Even along the coast, where Copiapoa occurs, summer UV Index commonly exceeds 11, classified by the World Health Organization as extreme risk.
Coastal Copiapoa are not exposed to this radiation unmodified. The Camanchaca moderates both visible and ultraviolet radiation while maintaining high atmospheric humidity, creating a light environment unlike that of inland deserts. Within the genus, persistent fog has a much greater influence on ultraviolet exposure than the relatively modest elevational range occupied by most populations. Elevation remains important for temperature, drainage, and fog interception, but fog frequency is the dominant factor shaping the coastal light environment.
Substrate further modifies these conditions. Pale granitic surfaces reflect more light while remaining comparatively cool, whereas dark volcanic substrates absorb and re-radiate heat, increasing thermal stress around the plant. These contrasting microclimates likely contribute to the striking differences in pigmentation, spination, and wax expression observed across the genus, although the relationship has not yet been tested directly at the locality level.
Together, intense sunlight, persistent fog, and substrate-driven microclimates have shaped the characteristic adaptations of Copiapoa. Reflective epicuticular wax (pruina), protective pigments, spine architecture, and CAM photosynthesis all reduce radiation stress while conserving water in one of Earth's most demanding environments. These are evolutionary adaptations, not short-term responses. A plant moved into brighter conditions cannot simply produce a thicker wax layer to compensate, which is why gradual acclimation remains essential in cultivation. The practical application of these light environments, including PAR targets, ultraviolet, and habitat replication, is discussed in the Cultivation section.

The fierce desert sun
*Art by AN Studio

Life here drinks the air
Camanchaca: The Fog That Sustains Life

Yet sunlight is only half the story. Without fog, Copiapoa would not exist.
Along Chile's northern coast, the cold Humboldt Current sustains vast banks of low marine stratocumulus clouds stabilized by the South Pacific Anticyclone. Persistent coastal winds carry this cool, moisture-laden air toward land. When the marine layer encounters the Coastal Range, it thickens and rises, forming the dense, dry fog known locally as Camanchaca. This drifting mist then moves inland through saddles and quebradas.
This fog is not mere atmosphere; it is the desert's lifeblood.
Camanchaca is classified as a dry fog, a phenomenon uncommon on Earth. Unlike typical fog, it carries a relatively low volume of liquid water and rarely produces sustained surface wetting or measurable soil moisture. The distinction matters: liquid water content and atmospheric humidity are different physical quantities. A fog can saturate the air with water vapor while delivering only minute amounts of liquid water to surfaces. Camanchaca does both, but the vapor pathway dominates. Life persists not through rainfall or surface wetting, but through the cumulative effect of repeated, minute atmospheric moisture inputs, sustained over hundreds of nights per year.
As the Camanchaca ascends the slopes, mountain ridges and concave faces intercept the densest flow of droplets. Micro-variations in elevation, aspect, and wind exposure create narrow ecological bands where fog persistence differs over only tens or hundreds of meters, exactly the scale at which Copiapoa populations segregate into distinct ecotypes.
The fog cycle and elevation limits
Although liquid water delivery remains low, nightly Camanchaca frequently drives relative humidity above 90% before dawn. The layer may rise more than 3,000 feet (approximately 1,000 m) along coastal slopes before thinning under intense sunlight. By late afternoon, humidity may drop to around 30%, only for the cycle to renew each evening, forming a reliable diurnal humidity pump that compensates for the desert's near-zero rainfall.
Across the region, fog penetration consistently weakens above about 800–1,100 m, creating the upper boundary of reliable fog-dependent ecosystems and marking the transition from mid-elevation fog belts to inland fog-shadow zones. This boundary is dictated by the marine inversion layer, a physical ceiling that traps moisture against the coast. Above this line, fog contribution drops sharply.
The inversion ceiling is not static. Direct microclimate measurements in Parque Nacional Pan de Azúcar documented that the marine inversion layer subsides in winter below the upper edge of the fog zone, exposing high-elevation sites to brief episodes of low humidity and elevated temperature even during the season normally associated with peak fog persistence (Thompson et al. 2003). The fog ceiling therefore moves on seasonal timescales, and its upper margin is where fog dependence is most exposed to year-to-year variability. Populations near the upper limit of Camanchaca penetration occupy a less reliable position than the near-permanent appearance of the fog layer would suggest.
🔴 For fog-dependent life, frequency matters more than depth: even minute droplet capture, repeated hundreds of nights per year, accumulates into vital hydration. Copiapoa cacti have evolved spines and surface structures that facilitate direct vapor capture, channeling moisture from spine to areole to stem.
Oasis de Neblina: The Isolated Fog Islands
Where the coastal Camanchaca meets land, it gives rise to unique fog-fed ecosystems known in Chilean literature as oasis de neblina (fog oases) and more broadly referred to as lomas. These develop along coastal foothills, cliffs, and escarpments, creating localized zones of biological productivity within an otherwise hyper-arid desert. The Atacama Desert has remained hyper-arid for millions of years, likely 15 to 25 million years or longer (Dunai et al. 2005; Rech et al. 2006), allowing these fog-dependent ecosystems to persist across immense spans of time. They support highly specialized plant and animal communities adapted to fog-derived moisture rather than regular rainfall.
🔴 Desert trick: The oasis de neblina are humidity engines, not wet islands
Mapped fog oases and geographic structure
These fog oases are not hypothetical or continuous features. Chilean floristic surveys and satellite analyses have identified and mapped more than 70 discrete oasis de neblina systems along the coastal Atacama, each functioning as an isolated fog-dependent ecological island (Rundel et al. 1991; Moat et al. 2021). Well-documented examples include Paposo-Taltal, Pan de Azúcar, Morro Moreno, Alto Patache, and Llanos de Challe, where topography consistently forces the Camanchaca to condense. These named fog oases correspond closely to the primary Geographic Anchors used on this site and provide the ecological framework within which Copiapoa populations persist, differentiate, and specialize.
Pan de Azúcar in particular has been the subject of multi-station microclimate monitoring documenting both the seasonal dynamics of the inversion layer and the elevational sorting of perennial species within individual quebradas (Thompson et al. 2003), providing one of the few primary datasets directly characterizing the climate conditions under which Copiapoa cinerea and associated species persist.
Where valleys and drainages (known locally as quebradas) cut through the Coastal Range, fog can penetrate inland along narrow corridors. These channels concentrate moisture well beyond the immediate coastline, allowing Copiapoa and other fog-dependent species to establish inland populations that could not normally survive on the exposed terrain between them. Many of the inland fog oases mapped across the Atacama Desert owe their existence to this channeling effect.
Coastal fog deserts develop vertically structured vegetation belts controlled by fog interception, elevation, and slope exposure, a pattern documented across the lomas ecosystems of Peru and northern Chile (Rundel et al. 1991; Dillon et al. 2003).
Plants such as Copiapoa, Tillandsia, Eulychnia, and other fog-dependent taxa have evolved mechanisms to exploit this vapor-based water source. In Copiapoa, spine architecture and epidermal structures modify the stem boundary layer and enhance fog interception, enabling persistence in landscapes that receive virtually no measurable precipitation.
Isolation and evolutionary consequences
Over long evolutionary timescales, populations within the genus Copiapoa have become finely tuned to distinct fog niches defined by elevation, slope orientation, substrate, and distance from the sea. These environmental gradients fragment populations into discrete fog oases that function as ecological islands, driving ecological differentiation and producing the remarkable diversity of form, color, and surface architecture observed across the genus. While this isolation reduces direct competition between populations, it also renders them highly sensitive to environmental disruption.
Independent floristic studies of non-cactus lineages in the Paposo–Botija coastal fog system confirm that oasis de neblina function as long-term biodiversity refugia structured by persistent Camanchaca, with sharp ecological boundaries and high vulnerability to declining coastal humidity and mining pressure (Ibáñez et al. 2022).
🔴 On the edge: Because these oases are separated by hyper-arid terrain, there is no meaningful “rescue effect.” If a unique evolutionary lineage at a specific site such as Quebrada Botija is lost, it is permanently erased.
Conservation threats and climate vulnerability
Today, these fog oases face increasing pressure. Shifts in fog frequency and inversion-layer behavior, likely linked to broader climatic change, combined with grazing, off-road activity, and illegal collecting, threaten the stability of systems that have persisted for millennia. Because most Copiapoa populations occupy small, fragmented habitats separated by extreme desert, even subtle changes in fog dynamics can produce long-term population collapse and irreversible local extinctions, as already observed in narrowly restricted, fog-dependent taxa such as Copiapoa solaris.
Some Atacama Desert soils contain extremely little organic matter, and early surveys underestimated how much microbial life could persist there. We now know that microbes can survive in Atacama Desert rocks and soils by exploiting mineral-bound moisture. Whether Copiapoa directly accesses these micro-reservoirs has not been experimentally demonstrated, but the root–rock interface and substrate chemistry likely influence stress, temperature, and moisture persistence in ways that shape long-term form.
In essence, the Atacama Desert is not defined by absence, but by adaptation. Its fog is both atmosphere and lifeline, an invisible river in the air that sustains an entire world of plants, from the humblest moss to the most enduring Copiapoa.
Camanchaca fog trapped against the Coastal Range (CR) defines the ecotype environment of Copiapoa
The Atacama: A Cool Desert Defined by Fog
Despite being one of the sunniest places on Earth, the coastal Atacama is classified as a cool desert. This is not incidental to Copiapoa survival. It is the precondition for it.
The interplay between the Camanchaca, the Humboldt Current, and constant coastal winds acts as a natural climate regulator, tempering the extremes that would otherwise occur at this latitude. Inland regions experience sharp day-night contrasts, with summer highs often reaching 86–95 °F (30–35 °C) and winter mornings dipping below 40 °F (4–5 °C). Along the coast, the fog's moderating influence is profound. Sites like Pan de Azúcar, Taltal, and Antofagasta maintain mild summer highs of 65–77 °F (18–25 °C) and winter lows near 50 °F (10–12 °C).
This thermal moderation matters because Copiapoa germination optima are unusually low for cacti, in some taxa near 15 °C rather than the 20-30 °C range typical of the family. A coastal climate that stays cool year-round is one of the few terrestrial environments where these germination requirements are met under natural rainfall, fog, and temperature conditions simultaneously. Even during occasional inland heat surges, the Camanchaca's nightly return reestablishes high humidity, resetting the desert's rhythm.
This constant cycle of condensation and dissipation, fog by night, sun by day, has made life possible for Copiapoa and countless other organisms that depend on vapor rather than rain.
🔴 In essence: The Atacama Desert is not defined by absence, but by adaptation. Its fog is both atmosphere and lifeline, an invisible river in the air that sustains an entire world of plants, from the humblest moss to the most enduring Copiapoa.
Ocean wind pushes cool Camanchaca fog into Atacama Desert coastal quebradas
Fog as Water: How Copiapoa Survives Extreme Aridity
In the hyper-arid Atacama Desert, where rainfall is negligible or absent for years at a time, Copiapoa persist by exploiting a water source that largely bypasses the soil: coastal fog. Although Camanchaca fog is “dry” by everyday standards, its frequency and persistence provide a dependable atmospheric moisture input capable of sustaining long-lived plants adapted to extreme aridity.
Engineering at the micro scale: fog capture and surface transport
Copiapoa spines are hydrophilic surfaces that intercept, collect, and directionally transport fog and dew droplets, a function additional to their shading and defensive roles rather than a replacement for them. A 2016 study, Hierarchical structures of cactus spines that aid in the directional movement of dew droplets in Philosophical Transactions of the Royal Society A (Malik et al.), documented this using time-lapse imaging, scanning electron microscopy, and a pilot tracer experiment. The plant was described there as Copiapoa cinerea var. haseltoniana, a taxon now referred to Copiapoa gigantea f. haseltoniana following the molecular treatment of Larridon et al.
Dew droplets were observed forming at spine tips and moving toward the base, including movement against gravity. Scanning electron microscopy revealed tapered microgrooves and a tip-to-base roughness gradient, structural features that drive surface water toward the areole. Evidence for movement past the areole into the stem came from a single preliminary experiment in which the spines of one areole of a juvenile plant were submerged in a suspension of 20-nanometer fluorescent microspheres for one hour, after which the excised section showed fluorescence in the cortex. The authors described this as a pilot result and noted that adult plants were untested.
Two surfaces are involved and should not be conflated. The spines themselves are hydrophilic, which is what lets them capture droplets and transport them to the areole. The epidermal pruina, by contrast, is hydrophobic and sheds water, keeping droplets mobile across the stem body and, under gravity, moving surface water down the plant toward the base.
Taken together, these results establish that Copiapoa spines capture fog and dew and directionally transport it to the areole. What becomes of that water at the areole is not resolved.
🔴 Technical insight (physical mechanism): Directional droplet movement along conical spines arises from differences in radius of curvature. These differences generate a Laplace pressure gradient that passively drives water toward the wider spine base at the areole, allowing efficient transport even against gravity.
A 2025 deuterium-labeling study (Huss et al., preprint) tested the inward step directly. Plants were exposed to isotopically labeled fog with the soil sealed, and cortex water was extracted afterward. In both Turbinicarpus alonsoi and Copiapoa humilis, cortex water showed no enrichment relative to unfogged controls. The authors also identified a suberized, water-impermeable layer at the spine base in Turbinicarpus and concluded that fog water runs down the plant surface to the soil for root uptake. The external runoff pathway proposed by Huss et al. has not yet been directly demonstrated in densely spined, heavily waxed coastal Copiapoa. Whether fog water reaching the areole subsequently enters the stem, remains on the plant surface, or is transferred to the soil under natural conditions remains unresolved for these taxa.
The role of areole wool
In Copiapoa, dense areole wool is positioned precisely at the point where spine-transported droplets arrive. While the water-retention function of areole wool has not yet been experimentally tested, its structure and placement may slow evaporation and increase the residence time of fog-derived moisture at the entry point into the stem. This potential role is consistent with the pronounced development of areole wool in fog-exposed taxa, but remains a hypothesis requiring direct study.
The nighttime recharge
Fog-driven hydration is most effective at night, when temperatures fall and relative humidity rises. Under these conditions, condensation on spines increases and boundary-layer humidity around the plant surface is elevated. These nighttime conditions coincide with CAM photosynthetic activity, during which stomatal opening is shifted away from daytime heat stress.
While the physiological coupling between CAM metabolism and fog-water transport has not been experimentally resolved, the repeated nightly cycle of condensation and dissipation creates regular opportunities for partial replenishment of internal water reserves before daytime solar stress resumes.
Roots: capturing surface moisture
Whether fog water first enters the stem or reaches the soil by surface runoff remains unresolved. Regardless of that pathway, Copiapoa possess a shallow, laterally spreading root system well adapted to capturing brief pulses of surface moisture from fog drip, dew, rare rainfall, and localized runoff.
Root excavations of Copiapoa cinerea ssp. columna-alba at Pan de Azúcar confirm this pattern. No roots were found deeper than about 8 cm (3.1 in), regardless of plant size (Gulmon et al. 1979, nomenclature historical). Although that study interpreted the root system in the context of rainfall, it predates the modern understanding of fog as the dominant source of moisture. The shallow architecture it documented is therefore more consistent with frequent surface wetting than with rare deep infiltration.
In addition to their shallow architecture, cactus roots are highly dynamic. Fine absorptive roots can develop rapidly following moisture events, greatly increasing the absorbing surface area before dying back during prolonged drought. These ephemeral roots allow rapid exploitation of short-lived water pulses without maintaining an extensive root system during dry periods (Dubrovsky and North 2002).
In the driest inland and high-elevation environments, certain Atacama soils can also retain small amounts of mineral-bound water through condensation at low humidity. Although direct uptake of this water by Copiapoa has not been demonstrated, the persistence of populations in fog-poor, rainless habitats suggests it may provide a limited supplementary source of moisture in fog-shadow and high-elevation environments.
Survival in a waterless world
Through the combined use of atmospheric fog capture and opportunistic root uptake, Copiapoa survive conditions that are lethal to most vascular plants. In an environment where rainfall is effectively absent, frequent but low-volume atmospheric moisture provides a dependable source of water while the shallow root system rapidly exploits brief pulses of surface moisture whenever they occur.
The fate of fog water after it reaches the areole remains one of the major unresolved questions in Copiapoa biology. Resolving it will require isotope-tracing experiments on mature coastal Copiapoa under natural nocturnal fog conditions, coupled with direct observation of water movement across the stem surface and into the root zone.
🔴 Fog logic: fog is the water source, and frequency matters more than volume.

Copiapoa spines channeling moisture to the areole. If water crosses into the stem is unresolved.
Beyond Water: Fog-Delivered Nutrients
Beyond water itself, fog delivers an additional resource. Beyond water itself, fog also delivers dissolved nutrients and organic material. Studies of fog-dependent ecosystems show that these atmospheric inputs influence plant growth, soil chemistry, and microbial activity.
Reciprocal transplant experiments on the fog-dependent bromeliad Tillandsia landbeckii demonstrate that atmospheric nutrients delivered by fog are a major driver of plant growth and elemental composition. Plants transplanted between fog zones gradually converged toward the nutrient content and growth patterns of local populations, reflecting differences in fog-borne nutrient availability. Nitrogen uptake closely tracked fog-derived nitrogen, whereas phosphorus remained more strongly regulated by internal growth demand.
Fog deposition also contributes nutrients and organic material to Atacama Desert surface soils and biological crusts, reinforcing the role of atmospheric inputs as a primary biogeochemical pathway in hyper-arid landscapes.
Interpretation
Copiapoa are rooted vascular cacti rather than atmospheric bromeliads, and direct equivalence should not be assumed. The bromeliad studies demonstrate ecosystem-level nutrient delivery via fog and plant-level nutrient dependence in fog-specialist taxa, but they do not test nutrient uptake pathways in Copiapoa.
However, Malik demonstrated transport along artificial cactus spine models and Opuntia spines, but not directly in Copiapoa. Whether either water or dissolved nutrients cross from the areole into the plant is subject to the same unresolved uptake question, so this remains hypothetical and has not been experimentally demonstrated.
The Water-Nutrient Paradox
Water and nutrients may follow different pathways
In most plants, water and nutrients are acquired together through the root system. In Copiapoa, the hyper-arid Atacama may partially separate these two processes.
Fog supplies frequent atmospheric moisture but wets only the uppermost millimeters of the substrate before evaporating. At the same time, the mineral-rich soils beneath the surface contain abundant phosphorus, potassium, calcium, magnesium, and other nutrients that remain largely immobilized because so little water moves through the soil profile. As a result, water may be relatively available at the surface during fog events while much of the substrate's nutrient reservoir remains physically inaccessible.
This limitation appears especially relevant to Copiapoa. Its shallow, laterally spreading roots are well adapted to capturing brief pulses of surface moisture rather than accessing deeper mineral reserves. Other plants with deeper or more aggressive root systems may exploit those reserves more effectively, suggesting that the constraint reflects the interaction between Copiapoa's root architecture and the extreme hydrology of the Atacama rather than an absolute lack of nutrients.
A working ecological model
This leads to a simple ecological hypothesis: water and nutrients may become partially decoupled in fog-dominated habitats. Atmospheric moisture is supplied frequently by fog, whereas nutrient acquisition depends on the relatively rare periods when surface wetting, microbial activity, and shallow root growth coincide. Growth may therefore be limited not by the total amount of nutrients present, but by how rarely those nutrients become biologically accessible.
This framework offers one possible explanation for the exceptionally slow growth of Copiapoa. Rather than experiencing prolonged periods of abundant water and nutrients, the plants may persist through thousands of small opportunities for resource acquisition, accumulating growth only gradually over decades. The model remains a working hypothesis and awaits direct experimental testing in habitat.
Summary
Taken together, these observations suggest that Copiapoa may be among the most atmosphere-dependent vascular plants known. Water, nutrients, and microbial activity all appear to be strongly linked to recurring fog events. This interpretation remains a hypothesis and will require targeted research for confirmation, including isotope tracing, nutrient budgeting under natural fog regimes, and direct measurement of root activity in fog-fed substrates.
Source Basis: Fog-delivered nutrient pathways and atmospheric deposition dynamics follow Ewing et al. (2008), González et al. (2011), Pinto et al. (2006), Weathers et al. (2010), and Fletcher et al. (2012). Interpretation of fog-driven microbial surface processes and shallow moisture penetration in hyper-arid substrates follows Azúa-Bustos et al. (2011) and Cowan et al. (2023). Discussion of phosphorus immobility and transport limitation in mineral systems follows Glenn et al. (1994), Föllmi (1996), and Yudovich & Ketris (2026). Full citations are provided on the Reference page.

Tilliandsia landbeckii, a Bromeliaceae airplant native to similar fog zones as Copiapoa cinerea
Microbial Ecology of Atacama Desert Soils

The Atacama Desert is often described as nearly lifeless, yet modern metagenomic studies reveal a surprisingly diverse microbial ecosystem beneath its barren surface. Even in the hyper-arid interior, far beyond the coastal fog belt, stress-adapted bacteria persist despite years that may pass between meaningful moisture events. Many of these microorganisms belong to previously undescribed lineages, indicating that the functional ecology of Atacama soils remains only partially understood.
This diversity does not imply continuous biological activity. Instead, most microorganisms appear to exist in a dormant state, becoming metabolically active only during brief periods when moisture becomes available. The result is not a biologically rich desert, but a sparse microbial reservoir capable of rapid activation when environmental conditions temporarily improve.
Although much of this community occupies the surrounding soil, association with plants creates localized refuges where moisture and organic carbon are more consistently available. This distinction between bulk soil and plant-associated microorganisms forms the basis of the rhizosphere and endosphere communities discussed below.
Plant-driven assembly of rhizosphere communities
Microbial communities in the Atacama are not simply passive inhabitants of the substrate. Plants actively shape the biological communities surrounding their roots through the release of root exudates that selectively recruit beneficial microorganisms.
Across a wide range of native Atacama plant species, the rhizosphere consistently contains greater abundances of nitrogen-fixing and plant growth-promoting bacteria than adjacent unvegetated soil. More recent work demonstrates that individual plant species assemble distinct microbial communities closely linked to local soil chemistry and root-derived metabolites. Rather than colonizing roots at random, microorganisms respond to chemical signals produced by the plant itself.
This selective recruitment establishes a dynamic ecological partnership. Beneficial organisms colonize root surfaces, some enter internal tissues as endophytes, and portions of these communities are later returned to the surrounding soil as roots grow and turn over. The rhizosphere therefore represents an actively maintained biological system rather than simply soil surrounding a root.
Fog-belt evidence
The most ecologically relevant evidence for this process comes from the coastal fog belt itself. Tillandsia landbeckii, a bromeliad that forms monospecific stands along the Coastal Cordillera between approximately 900 and 1,300 m elevation, occupies the same hyper-arid fog corridor as many Copiapoa populations and depends almost entirely on fog for water input.
A study of bacterial communities associated with Tillandsia landbeckii demonstrated that the plants hosted microbial assemblages distinct from the surrounding substrate and that these communities varied geographically and seasonally with fog availability. Rather than representing passive deposition of airborne microorganisms, the evidence suggests that fog-dependent plants actively maintain structured, environmentally responsive microbiomes. (Hakobyan et al. 2023)
Among currently studied systems, Tillandsia represents one of the closest ecological analogues for understanding microbial dynamics in coastal Copiapoa habitats. The evidence suggests that fog functions not only as a water source for the host plant, but also as a regulator of the associated microbial community.
Endophytic associations in desert cacti
The relationship between desert plants and their microbial partners extends beyond the external rhizosphere. Research on desert cacti indicates that endophytic bacteria are not merely incidental inhabitants but may contribute functionally to host establishment and growth under extreme aridity, with the size of that contribution depending heavily on substrate nutrient availability.
Functional dependence in cardon
The strongest functional evidence for the importance of endophytic bacteria comes from the giant cardon cactus (Pachycereus pringlei). Endophytes isolated from roots and seeds were shown to fix atmospheric nitrogen, solubilize phosphate, weather mineral substrates, and release organic acids capable of mobilizing otherwise inaccessible nutrients.
Their ecological importance was demonstrated experimentally. Removing native endophytes had little effect on seed germination but significantly reduced seedling growth and root development, while reintroducing the bacteria restored normal vigor. Under nutrient-limited mineral conditions, the microbial community became an essential component of early plant establishment rather than simply a beneficial addition.
Seed-borne transmission
Evidence from both Pachycereus pringlei and Mammillaria fraileana indicates that at least part of the endophytic community is transmitted through seed rather than acquired entirely from the surrounding environment. Endophytes have been detected within embryos, vascular tissues, and developing seedlings, suggesting that each new generation begins life with an inherited microbial community that is later reinforced through rhizosphere recruitment.
Substrate-dependent benefit
The benefits provided by endophytes are not universal but depend strongly on environmental conditions. In Mammillaria fraileana, inoculation improved nutrient acquisition and enhanced CAM activity only when seedlings were grown on native mineral substrates. This suggests that microbial partners become most valuable where nutrient availability is constrained by the substrate itself, conditions that closely resemble those experienced by many habitat-grown Copiapoa.
Two moisture regimes
No equivalent metagenomic work has yet been conducted directly on Copiapoa. Rhizosphere and endosphere communities associated with the genus therefore remain uncharacterized. Nevertheless, the combined evidence from Atacama soil surveys, fog-belt microbiome studies, cactus endophyte research, and rhizosphere chemistry allows a plausible ecological framework to be proposed.
The same moisture limitation that restricts nutrient mobility in hyper-arid substrates also constrains the activity of free-living soil microorganisms. In the coastal fog belt, rhizosphere organisms in the surrounding substrate are likely subject to the same episodic activation cycle as the broader ecosystem, becoming metabolically active only during brief periods when fog condensation wets the shallow substrate (Azua-Bustos et al. 2011; Connon et al. 2007). As the substrate dries, most external microbial processes likely return to dormancy.
The plant interior represents a fundamentally different hydrological environment. Endophytic microorganisms colonizing root cortex tissue and internal structures operate within tissues hydrated by water already captured and retained by the plant. In succulents with persistent water storage capacity, this internal environment may remain hydrated long after surrounding soil has dried.
Microbial activity within the plant body may therefore be partially decoupled from the external fog cycle governing the substrate. Endophytic partners could continue contributing low-level physiological services, including nitrogen fixation, phosphorus mobilization, mineral weathering, and stress-related signaling, between external wetting events by utilizing plant-derived water and carbon resources (Puente et al. 2009a; Lopez et al. 2011).
The result is a system operating across two distinct moisture regimes: pulsed activity in the external rhizosphere during fog events, and potentially more sustained activity within hydrated plant tissues between them. The substrate briefly wets, activating free-living microbial exchange and nutrient mobilization. Inside the plant, endophytic communities may continue functioning for longer periods using the plant itself as a buffered microenvironment.
Implications for understanding Copiapoa
For a genus with growth rates measured in millimeters per decade, even modest microbial contributions sustained over long periods may become physiologically significant. The substrate environment in which Copiapoa evolved was not biologically inert, but part of a sparse, fog-regulated, microbially structured ecosystem in which stress-adapted organisms operated both around and within the plant body.
External rhizosphere communities likely activate during fog pulses, while endophytic communities may continue functioning between those events using plant-retained water. Over decades or centuries, each individual plant may gradually shape and reinforce a localized microbial system through repeated cycles of root exudation, microbial recruitment, endophytic colonization, and reintroduction into surrounding substrate.
This framework also has implications for understanding why cultivated Copiapoa often diverge from habitat morphology. In cultivation, seedlings are commonly established in sterile or biologically simplified substrates and develop without prolonged exposure to the fog-belt microbial communities present in habitat. The feedback loop that would normally assemble a habitat-like rhizosphere is therefore unlikely to establish fully under conventional cultivation conditions.
If cultivated plants lack comparable endophytic partners, they may rely more heavily on freely available dissolved nutrients in the substrate solution. Under horticultural conditions, this could contribute to accelerated growth rates and metabolic profiles that differ substantially from those maintained in habitat environments. Such shifts may occur alongside the more familiar effects of excess water, nutrient availability, reduced environmental stress, and altered light conditions, potentially contributing to softer structure, weaker spination, and diminished pruina development.
Source Basis: Ecological interpretation in this section is synthesized from published research on Atacama Desert microbial ecology, rhizosphere assembly, fog-dependent plant systems, and desert cactus endophytes. Hypolithic fog-supported microbial communities follow Azúa-Bustos et al. (2011). Hyper-arid soil microbial diversity and actinobacterial persistence follow Connon et al. (2007) and Idris et al. (2017). Rhizosphere enrichment and plant-driven microbial recruitment in Atacama plants follow Eshel et al. (2021), Fortt et al. (2025), and Dussarrat et al. (2025). Fog-belt microbial systems associated with Tillandsia landbeckii follow Hakobyan et al. (2023).
Endophytic rock-weathering, nutrient mobilization, and seed-borne transmission in Pachycereus pringlei follow Puente et al. (2009a, 2009b). Endophyte detection, vertical transmission, and substrate-dependent growth promotion in Mammillaria fraileana follow Lopez et al. (2011, 2012). Extension to Copiapoa, for which no comparable endophyte data exist, represents evidence-based ecological inference rather than experimentally validated cultivation protocol. Full citations on the References page.
Pruina (not Farina) and Apical Wool: Copiapoa’s Defenses
In the fog-influenced coastal deserts of northern Chile, one of the driest and most light-intense environments on Earth, Copiapoa cacti have evolved two of their most visually distinctive protective features: the silvery veil of pruina (epicuticular wax) coating the epidermis, and the dense apical wool associated with flowering and reproduction. Together, these structures regulate light, temperature, and moisture in habitats defined by extreme solar radiation and near-total absence of rainfall.
Pruina, not “Farina”: correct terminology for Copiapoa wax
The chalky white or bluish coating seen on many Copiapoa is properly termed pruina, referring to a layer of epicuticular wax composed of microcrystalline lipids deposited on the plant surface. This wax bloom functions as a protective interface, increasing reflectivity (albedo), reducing ultraviolet and thermal stress, and moderating boundary-layer humidity at the epidermis.
The term “farina” is widely used in horticulture as a colloquial descriptor for powdery surface coatings on plants, but it is imprecise and not anatomically specific. In botanical and physiological literature, the structure present on Copiapoa and other wax-coated succulents is consistently described as epicuticular wax or pruina. Using the correct term avoids confusion with other types of surface powders, fungal residues, or particulate deposits that may appear superficially similar but differ in structure and function.
For clarity and scientific consistency, this site uses pruina (epicuticular wax) when referring to the natural wax bloom on Copiapoa epidermis.
Pruina: the Living Mirror
Pruina is a dense layer of microscopic epicuticular wax crystals secreted by the epidermis. Within the Camanchaca fog belt, this wax coating performs several critical functions shaped by the region’s unusual light environment. Epicuticular waxes form hierarchically structured micro- and nanocrystalline surfaces that strongly influence reflectance, light scattering, wettability, and thermal behavior, functioning as the primary interface between the plant and its environment (Barthlott et al., 2017).
Pruina reflects high-energy ultraviolet radiation that penetrates coastal fog while simultaneously scattering visible light within the epidermal layers. This scattering improves internal light distribution under diffuse illumination, a persistent condition in fog-dominated habitats. At the same time, the wax layer lowers stem surface temperature, reducing heat load during periods of intense solar exposure.
The wax crystals also create a highly hydrophobic surface. Fog droplets bead and roll off rapidly, limiting prolonged surface wetting while still allowing atmospheric moisture to interact with the plant surface. In addition, pruina reduces cuticular transpiration during rainless intervals that may last for years or decades.
The structural classification of Copiapoa wax helps explain what the pruina layer actually is. Barthlott et al. (1998), in a comprehensive SEM survey of epicuticular waxes across more than 13,000 plant species, classified the wax on Copiapoa cinerea as a "fissured layer": a thick, crusty covering that fractures into terraced plates as the epidermis expands. This cracking occurs because epidermal expansion and wax deposition proceed at different rates. On any plant producing this type of wax, the visible crust reflects the balance between the plant's inherited capacity for wax deposition and the rate at which new epidermis is produced. When the epidermis expands faster than new wax accumulates, the crust becomes thinner and more extensively fissured. When growth is slower, the same wax-producing capacity maintains a thicker, more continuous layer.
In coastal Copiapoa, that balance is striking. Fog-belt populations receive the most consistent moisture in the genus and ultimately produce the largest plants Copiapoa attains. Their epidermis expands at rates that would thin or fracture the wax crust on plants with lower secretion capacity. Yet coastal plants maintain the thickest pruina in the genus. The inherited capacity for wax deposition is high enough that even relatively vigorous growth does not overwhelm the wax layer.
Ecological tradeoffs and energy investment
The capacity for heavy wax deposition is genetically structured, though its visible expression still interacts with environmental conditions and growth rate. It requires energy that plants in drier, less fog-buffered zones cannot afford to spend. Coastal populations evolved this investment because the return justified the cost. In the fog belt, where direct PAR is chronically suppressed by persistent stratus and a substantial proportion of available light arrives as diffuse radiation, a thick wax crust that scatters incoming light internally and reflects damaging ultraviolet radiation is not a luxury. It is the mechanism by which the plant makes productive use of the limited photosynthetic light reaching its surface. Higher-zone plants, receiving more direct PAR but less moisture, face a different energy budget. They cannot sustain the same rate of
wax deposition and instead allocate resources toward other strategies suited to their conditions.
Scientific basis for the “Living Mirror” model
The functional interpretation of Copiapoa pruina is well supported by broader research on plant epicuticular waxes.
The structural foundation for this model begins with the wax itself. Barthlott et al. (1998), in the standard classification of plant epicuticular waxes based on SEM analysis of over 13,000 species, identified the wax on Copiapoa cinerea as a fissured layer, one of 23 defined wax types. Fissured layers are thick, crusty coverings characteristic of succulents, built by continuous deposition and fractured into terraced plates as the underlying epidermis expands.
This classification establishes that Copiapoa pruina is not a thin film or a loose powder but a massive, structured crust. In coastal ecotypes, the genetic capacity for wax secretion is high enough to build and maintain this crust even on the fastest-growing plants in the genus, an adaptation that only the fog belt's moisture budget makes metabolically viable.
Shepherd and Griffiths (2006) demonstrated that filamentous and rodlet-type wax crystals, structurally comparable to those reported from coastal Copiapoa, strongly influence surface optical behavior. These waxes increase reflectance, scatter visible light, and modify internal light distribution within the epidermis under stress conditions.
Work summarized by Koch and Ensikat (2008) further shows that microcrystalline wax layers reduce heat load and act as both physical and optical barriers, protecting underlying tissues from ultraviolet damage and thermal stress.
More recent structural analyses of plant epidermal micro- and nanostructures confirm that epicuticular waxes act as efficient light scatterers, modifying reflectance, internal light distribution, and surface temperature independently of pigmentation. These properties arise from the hierarchical organization of wax crystals and cuticle texture rather than from chemical coloration.
Taken together, these findings provide a strong scientific foundation for describing pruina as a functional “living mirror”: a wax layer that limits harmful radiation while improving internal light distribution under the fog-dominated conditions characteristic of coastal Copiapoa habitats.
🔴 More than UV protection: Pruina is often presented as little more than protection against ultraviolet radiation. In Copiapoa, however, its geographic distribution is more consistent with a broader ecological role. The heaviest wax occurs not in the driest or sunniest habitats, but in the coastal fog belt, where it likely regulates light across alternating periods of dense fog and intense sunshine.
Evidence from cultivation
Some of the most intensely white coastal Copiapoa ever cultivated were produced not in Chile or California, but in cool, diffuse-light European greenhouses.
Well-documented examples include large coastal plants grown by Roger Kropf in Switzerland, exceptionally pruinose Copiapoa gigantea cultivated by Heinz Hoock and others in Germany, and snow-white colonies maintained at the Zürich Succulent Collection and at Specks nursery. These plants often equaled or exceeded habitat specimens in pruina thickness, not because European conditions stimulated greater wax deposition, but because cool temperatures and diffuse light kept epidermal expansion relatively slow. The coastal ecotype's inherited capacity for wax deposition could therefore maintain a thick, continuous wax crust rather than being spread across rapidly expanding tissue.
Inland and high-montane ecotypes grown under the same conditions remained comparatively green, demonstrating that favorable cultivation alone cannot produce heavy pruina. The difference lies in inherited wax-deposition capacity. The cool, diffuse conditions of European greenhouses limited epidermal expansion, allowing coastal genotypes to maximize the expression of their inherited wax-producing phenotype.
The geography of wax: why pruina expression varies
While the potential to produce epicuticular wax is a hallmark of the genus, its expression is not uniform. Pruina density follows stable, genetically inherited ecotypes rather than short-term environmental responses. Because the metabolic cost of wax deposition is high, the degree of "whiteness" seen in a population serves as a morphological signature of its specific habitat.
In the coastal fog belt, persistent fog provides the moisture budget that supports the heaviest wax expression while creating a highly variable light environment. The resulting thick wax layer both moderates excess radiation during clear conditions and improves the utilization of diffuse light beneath fog. Inland fog-shadow habitats and high-elevation populations experience fundamentally different moisture and light regimes, favoring alternative epidermal strategies. The result is a predictable ecological gradient, a cline of physiological adaptation that remains stable even when plants are moved into cultivation.
Source Basis: Epicuticular wax classification and structural morphology follow Barthlott et al. (1998). Functional properties of plant surface waxes follow Barthlott et al. (2017), Shepherd and Griffiths (2006), Koch and Ensikat (2008), and Riglet et al. (2021). Fog climatology follows Jung et al. (2020) and Böhm et al. (2021). Cultivation observations are from published and documented collections. Full citations are on the Reference page.

Light-scattering pruina formed by epicuticular wax crystals in coastal fog belt (Zone 1) Copiapoa
Apical Wool: Protection at the Growing Point
Copiapoa do not form true cephalia in the botanical sense. Instead, they develop dense apical wool associated with flowering and reproduction.
A true cephalium, as seen in genera such as Melocactus, represents a permanent developmental transition in which vegetative stem growth ceases and a dedicated reproductive structure forms. In Copiapoa, apical wool remains an extension of the active vegetative meristem. Stem growth continues, and the wool does not represent a terminal or irreversible phase.
The wool consists of elongated hairs and bristles produced by areoles near the growing apex. It occurs across the genus in both columnar and globular forms. In mature plants, this woolly crown functions as a protective zone over the meristem and developing flowers.
Functional role: shield and reproductive microclimate
In tall-growing taxa such as Copiapoa gigantea and in coastal forms within the Copiapoa cinerea complex, apical wool becomes especially dense at maturity. It serves two primary functions.
➤ First, it acts as a solar and thermal buffer, diffusing incoming radiation and reducing direct exposure of sensitive meristematic tissue. Many plants exhibit a consistent northward orientation of the woolly apex, aligning with the sun’s path in the Southern Hemisphere. This orientation reduces peak irradiance while allowing gradual warming.
➤ Second, the wool creates a stable reproductive microclimate. Flowers and fruits emerge from within the wool, where humidity is retained, temperature fluctuations are dampened, and developing tissues are protected from desiccation and ultraviolet stress.
Ecological grouping of apical wool expression (illustrative)
Apical wool expression follows stable ecological and locality-linked patterns, not taxonomic rank. The following groupings are descriptive only and summarize recurring field-observed expressions:
- Golden to orange-tinted wool
Copiapoa gigantea
High visual contrast against pale epidermis in large-bodied coastal and mid-elevation populations. - Narrow, pale wool
Copiapoa cinerea f. columna-alba
Low contrast, blending with dense pruina in coastal fog belt ecotypes. - Dense white to beige wool
Copiapoa dealbata; Copiapoa cinerea f. krainziana
Compact wool associated with slow-growing, stress-adapted populations.
These groupings are ecological descriptors only and do not imply species or subspecific rank.
Terminology clarification: “pseudo-cephalia”
Historical literature, particularly mid-20th-century treatments, occasionally applied the term pseudo-cephalium to intensified apical wool in Copiapoa. This terminology reflected an attempt to describe conspicuous flowering zones using the conceptual framework available at the time.
Modern botanical interpretation does not recognize these structures as cephalia or transitional organs. They are understood as localized intensifications of the vegetative meristem associated with flowering, without a permanent shift in growth mode.
Source Basis: Pruina function and optical properties follow Shepherd & Griffiths (2006), Koch & Ensikat (2008), and Barthlott et al. (2017). Pruina expression across ecotypes draws on documented habitat photography and long-term cultivation records. Full citations are on the Reference page.
From Crown to Provenance
The morphological features described above (pruina density, spine structure, apical wool development) are not random variations. They are expressions of specific environmental pressures: fog frequency, elevation, substrate, and UV exposure. Tracking these pressures is the role of locality data and field numbers.
A plant originating from a coastal Zone 1 locality typically exhibits dense wax and wool adapted to diffuse light and persistent atmospheric moisture. A plant from a Zone 3 inland fog-shadow locality reflects adaptation to extreme aridity, mineral buffering, and high ultraviolet exposure.
Interpreting apical wool alongside provenance data is essential for maintaining habitat-appropriate form in cultivation. Guidance on preventing wool reduction, pruina loss, and inappropriate growth responses is provided in the Generalist vs. Specialist Care section.
From Habitat to Cultivation: Tracking Copiapoa Origins
Field Number and Locality Information
Understanding a Copiapoa’s origins begins long before it enters cultivation. In habitat, each population is shaped by a repeatable set of environmental variables: fog frequency, which modulates light intensity, diffusion, and spectral composition, thereby influencing effective PAR exposure, elevation, slope exposure, substrate chemistry, and ultraviolet radiation. These factors leave a lasting imprint on plant structure and physiology and continue to influence growth form and physiological response long after removal from habitat.
Together, these variables define a plant’s ecotype zone, which determines its optimal light levels, watering frequency, temperature tolerance, and long-term care requirements. Cultivation that ignores ecological origin inevitably leads to stress, decline, or loss.
Locality shapes more than visible morphology. Field and metagenomic studies of Copiapoa solaris rhizosphere communities document that microbial associations differ meaningfully across populations separated by as little as 25 km, tracking local humidity gradients and thermal load. The microbial community a plant carries from habitat is part of its ecological identity, not an incidental feature. Cultivation practices divorced from ecological origin may affect this dimension of a plant's biology in ways that are invisible until long-term stress or decline makes them apparent.
For this reason, locality information is the most biologically meaningful data point associated with any Copiapoa, and it remains informative even in the absence of a formal field number. Locality identifies where a plant or seed originated and, by extension, which ecotype zone it belongs to. A field number is simply a collector’s unique identifier that anchors that locality data to a specific population or seed harvest.
In some cases, plants circulate with complete locality descriptions but without an official field number. When locality data is accurate and detailed, it remains fully sufficient to identify the correct ecotype and cultivate the plant successfully.
What are Field Numbers and Locality Data?
Field Number
A unique alphanumeric code (for example PV2146, RH2087, KK1523) assigned by a collector to a specific plant, population, or seed harvest.
Locality Information
The descriptive data associated with a field number, or recorded independently when no field number exists. This may include:
- GPS coordinates (exact or approximate)
- Elevation
- Slope aspect
- Distance from the coast
- Substrate type
- Associated vegetation
- Fog exposure or microclimate notes
Together, or locality alone when a field number is missing, these function as the plant’s passport and ecological fingerprint, establishing provenance and identifying which ecotype zone the plant belongs to: coastal fog belt, transitional fog belt, inland fog-shadow, or high montane.
Copiapoa with field number tags
Field Number Example (Applied)
- Field Number: PV2146
- Collector: Petr Pavelka (son)
- Species (legacy name): Copiapoa columna-alba (within the cinerea complex)
- Locality: Chile, northeast of Esmeralda; south-facing slope above the fog inversion
- Ecotype interpretation: Transitional fog zone near the upper coastal inversion boundary.
Locality without a Field Number
Copiapoa cinerea (complex) → San Ramón Valley, hillside above Taltal, south-facing slope at approximately 800 m (no field number; locality documented by grower at acquisition; legacy name C. krainziana)
This illustrates that detailed locality description without a formal field number retains full conservation and cultivation value, provided the geographic and ecological data are specific enough to identify the ecotype zone and population context.
Why locality data matters more than the species name alone
Two plants labeled Copiapoa cinerea can belong to entirely different ecotype zones and require radically different care.
➤ Coastal fog belt origin (for example Taltal-sur or Paposo) plants originating from persistent coastal fog zones typically show:
- Heavy chalk-white epicuticular wax (pruina) adapted to diffuse, high-UV coastal light
- Adaptation to moderated but frequent fog-filtered irradiance rather than extreme inland solar intensity
- Tolerance for relatively regular, light hydration pulses reflecting repeated fog deposition rather than rare deep rainfall events
➤ An inland fog-shadow or high montane origin (often above ~1,000–1,100 m) typically produces:
- Darker or bronze epidermis
- Full sun with extreme ultraviolet exposure
- Very infrequent, deep watering events
Without accurate locality data, growers risk applying inappropriate care that can stress or permanently damage mature specimens.
Role in cultivation and conservation
Locality data, and field numbers when available, are essential for:
- Matching plants with correct ecotype-specific care
- Responsible breeding within the same population or compatible zones
- Preventing undocumented hybridization and genetic erosion
- Supporting conservation through accurate provenance tracking
🔴 Full locality information is a Copiapoa’s true birth certificate: It tells you not just what the plant is, but which version of that species you are growing, and how to keep it habitat correct.
Addressing Incorrect Legacy Labeling
Correcting the record without erasing history
Much of the confusion surrounding Copiapoa stems from legacy labeling practices that elevated local morphologies to species rank before ecological context and molecular data were available. As a result, many cultivated plants retain outdated names that no longer reflect current taxonomy, even when provenance is well documented.
Correcting these labels does not mean discarding historical information.
Primary vs. supplementary data
Original labels, collector names, and legacy identifications are primary data, not annotations. Once removed or overwritten, the original evidentiary state cannot be reconstructed.
Documentation loss rarely occurs all at once. It accumulates through small, well-intentioned changes: a shortened locality, a removed collector number, a replaced name without a note. Over time, provenance and scientific value are irretrievably degraded.
This is not a theoretical concern. Larridon et al. 2014 demonstrate that undocumented or mixed ex situ collections fail to preserve population-level genetic structure, limiting their usefulness for conservation, research, and future reintroduction efforts.
The solution is to separate taxonomy from provenance:
- Taxonomy: provisional, revisable
- Provenance: permanent, non-negotiable
Best practice is therefore not to replace an old name with a new one in isolation, but to reframe the label to communicate both current understanding and historical context.
Recommended labeling format
Species (modern taxonomy) + precise locality and/or field reference,
with legacy names retained parenthetically or in notes.
Examples:
Copiapoa gigantea → Paposo area, coastal slope (ex FR 208; formerly labeled C. columna-alba)
Copiapoa cinerea (complex) → North-East of Esmeralda, Chile (PV 2146; legacy name C. columna-alba)
This approach:
- Aligns plants with current taxonomic consensus
- Preserves historical labels and collector data
- Retains ecotype and habitat context essential for cultivation
- Keeps older literature, seed lists, and photographs interpretable
- Allows future reinterpretation under improved frameworks
Legacy names such as columna-alba, krainziana, melanohystrix, or haseltoniana should not be erased. Their value lies in documenting historical interpretation, morphology, and locality, not in asserting current species boundaries.
Because many historically documented plants exist only in private collections, growers function as de facto custodians of irreplaceable data. Digital records are valuable supplements, but they do not replace permanent physical labels, which remain the only data guaranteed to stay with the plant over decades of cultivation.
🔴 Principle: Update the name. Never erase the history.
Source Basis: Provenance tracking and population integrity follow Larridon et al. (2015) and Davis & Pillet (2023). Full citations are on the Reference page.