Ecotype, Hybridization, and Variation in Copiapoa
Three Sources of Variation
This guide separates three sources of variation that are often confused: natural hybridization, horticultural hybridization, and ecotype-driven morphology. Natural hybridization occurs in Copiapoa, but it is usually local and uncommon. Cultivation can produce undocumented hybrids much more easily because natural barriers are removed. Most wild variation, however, is best understood through fog regime, elevation, substrate, radiation, and long-term ecological specialization.
Hybridization: Real, Local, and Often Misunderstood
Natural hybridization in habitat
For millions of years, Copiapoa species have evolved within the fog-fed, geographically fragmented landscapes of Chile’s Atacama Desert. Populations are often separated by broad stretches of barren terrain, steep topography, and sharply different local climates. Over evolutionary time, this isolation has encouraged ecological specialization while helping preserve distinct genetic lineages.
Natural hybridization does occur in Copiapoa, but it appears to be uncommon and geographically limited. Where the ranges of closely related species overlap, shared pollinators and synchronized flowering can occasionally produce intermediate plants. These cases are best documented in narrow contact zones within parts of the Copiapoa humilis and Copiapoa solaris complexes.
These examples are important because they show that interspecific hybridization is possible in the wild. However, they do not support the idea that most visible variation in Copiapoa is the result of widespread hybrid mixing. In habitat, hybridization remains local, occasional, and constrained by the same environmental barriers that shape the genus as a whole.
Why natural hybridization remains localized
In the wild, both pollen and seed movement are limited.
Copiapoa flowers are primarily insect-pollinated and lack the traits associated with long-distance wind or vertebrate pollination. Among the documented floral visitors are jewel beetles of the genus Ectinogonia. These beetles feed on Copiapoa flowers and can transfer pollen between nearby plants, with adults of Ectinogonia superba recorded completing their larval cycle within Copiapoa humilis and Copiapoa cinerea columna-alba at coastal Atacama Desert localities (Pineda and Mondaca 2020). These pollinators operate at the scale of local plant groups, not across broad landscapes. As a result, pollen movement is usually confined to nearby colonies or narrow zones where compatible species grow close together.
Seed dispersal is similarly restricted. Copiapoa seeds are relatively heavy and usually fall close to the parent plant. Ants may move some seeds short distances after being attracted to nutritive seed tissues, but this still reinforces local rather than long-distance dispersal. Although the Atacama Desert is windy, Copiapoa pollen and seeds do not show adaptations for long-distance wind transport.
Together, these factors keep gene flow highly localized. Even where compatible species occur near one another, natural hybridization usually remains confined to small contact zones rather than spreading across entire regions.
Historically, many intermediate forms within the Copiapoa cinerea complex were interpreted as hybrids. Modern ecological and molecular evidence points to a different explanation. Much of this variation is better understood as long-term ecotypic differentiation and phenotypic plasticity shaped by elevation, fog frequency, substrate, and thermal regime.
These plants are not recent blends. They are populations shaped over thousands to hundreds of thousands of years by stable environmental pressures. Their differences reflect local adaptation within continuous evolutionary lineages, not hybrid swarms.
Cultivation removes natural barriers
Cultivation changes the situation completely.
In habitat, distance, pollinator behavior, flowering overlap, and seed dispersal all restrict hybridization. In cultivation, those barriers are removed. Plants from different regions, species, or ecotypes may grow side by side and flower at the same time. They may be hand-pollinated intentionally, or they may cross-pollinate unintentionally in mixed collections.
Cultivated hybrids can be beautiful, vigorous, and horticulturally valuable. Well-documented hybrid breeding has produced respected lines in Japan, Europe, and the United States, especially within the cinerea, humilis, and solaris groups. When clearly labeled, these plants are legitimate horticultural creations.
The problem is not hybridization itself. The problem is the loss of accurate lineage information.
Undocumented hybrids can circulate for years as “pure species,” blurring taxonomic clarity and contaminating seed lines. Because Copiapoa grow slowly, hybrid ancestry may remain hidden for decades before certain traits become visible. This delayed expression, often described as cryptic introgression, makes undocumented hybridization especially difficult to detect once it enters cultivated seed pools.
For collectors, researchers, and conservationists, accurate records are therefore essential.
Labeling, provenance, and habitat seed
Hybrids should always be labeled clearly. A plant produced from Copiapoa cinerea and Copiapoa humilis, for example, should be written as:
Copiapoa cinerea × Copiapoa humilis
In hybrid notation, the seed parent is listed first, followed by the pollen parent. This convention preserves useful information about the cross and prevents confusion later.
Some naturally occurring hybrids are widely recognized among growers, but they remain hybrids by definition. A plant may be attractive, stable, and well known in cultivation while still being unsuitable as a representative of a pure wild lineage.
Accurate provenance and locality data are just as important. Plants without reliable origin information are often unsuitable for taxonomic, ecological, or conservation study, no matter how closely they resemble a published name.
Seeds described as coming “from habitat plants” are not necessarily habitat-collected seeds. If a wild-collected parent plant flowers in cultivation alongside other Copiapoa species, open pollination can produce undocumented hybrid seed. This is one of the most common sources of cryptic hybridization in cultivated Copiapoa. A seed batch is only equivalent to habitat seed if pollination occurred in habitat or if the cultivated pollination history is clearly documented. Without that information, the offspring should not be treated as verified wild-lineage material.
With proper records, hybrids can be appreciated for their horticultural value without compromising ecological or evolutionary understanding.
🔴 Principle: Hybridization may blur boundaries in nature, but records must remain precise.
When to suspect undocumented hybridization
Suspect hybrid origin when a plant shows trait combinations not known from documented wild populations. Warning signs include:
- intermediate spine structure combined with atypical epicuticular wax
- growth habits outside the known range of the relevant ecotype
- unusual vigor paired with uncertain provenance
- conflicting traits from geographically or ecologically separate lineages
By contrast, plants from well-documented localities that show variation consistent with their ecotype zone are more likely expressing environmental adaptation than hybrid ancestry.
Serious collectors should compare uncertain plants against documented habitat specimens, field photography, and well-curated institutional collections. References such as the Sarnes monograph, historical field documentation, and verified locality material provide the baseline needed to distinguish normal ecotypic variation from anomalous traits.
When origin is uncertain and traits appear inconsistent, the plant should be labeled conservatively as “uncertain origin” or “suspected hybrid” rather than assigned confidently to a pure species.
From hybrid misconception to ecotype interpretation
Once the limited role of natural hybridization is understood, the larger question becomes clearer: why can genetically coherent plants look radically different within the same named lineage?
Recent macroevolutionary analyses support an ecological answer, showing that cactus diversification is shaped by interacting drivers including geographic distribution, plant morphology, soil composition, climate, and topography (Thompson et al. 2024). In Copiapoa, these forces are especially visible because the Atacama Desert presents extreme and persistent environmental gradients over short distances.
Across much of the genus, visible variation appears to be driven primarily by ecological factors rather than recent genealogical mixing. Fog regime, elevation, substrate, radiation, and long-term habitat stability shape how plants express form, color, spination, wax, and growth habit over evolutionary time.
Source Basis: Spatially constrained gene flow and limited natural hybridization follow Larridon et al. (2014, 2015). Hybridization risk in cultivation and provenance management practices reflect Sarnes (2025) and Davis & Pillet (2023). Full citations are on the Reference page.

Japanese cultivated Copiapoa cinerea hybrid (horticultural cross)

Hostile substrate at Pan de Azucar strongly favors Copiapoa
The Ecotype Model: Why Wild Plants Look Different
Variation is not random
Copiapoa do not vary randomly. Their forms reflect the climate bands and exposed geological mosaic of the Atacama Desert.
The most dramatic differences in Copiapoa span the full spectrum: snow-white versus jet-black bodies, soft water-rich ribs versus hardened bronze forms, and long fog-intercepting spines versus short upright spines. These contrasts are not usually the product of different species or widespread hybridization. They arise from long-term adaptation within related lineages as individual populations respond to local environmental pressures over thousands of years.
Across the Atacama Desert, predictable combinations of fog exposure, solar radiation, temperature extremes, substrate chemistry, and slope orientation repeat along the coast and across elevation bands (Moat et al. 2021). When Copiapoa populations establish within similar environmental corridors, they often develop similar growth forms, even when separated by hundreds of kilometers.
Repeated environmental structures produce repeated selective pressures, and therefore repeated plant forms. This explains why distant populations can appear nearly identical, while plants bearing the same name may behave very differently in cultivation. Accurate interpretation therefore depends on environmental history rather than surface resemblance alone.
This relationship between form and environment is not only observable at landscape scale, but has been directly measured in the field. Mooney, Weisser & Gulmon (1977) conducted one of the earliest ecophysiological field studies of the genus, examining thermal relations and environmental adaptations in plants near Paposo, treated in the paper as Copiapoa haseltoniana but now better understood as a coastal form within the broader Copiapoa gigantea lineage. Their findings showed that body orientation, reflective pruina, epidermal surface properties, and spine architecture operate as an integrated system for managing radiation and temperature in a coastal, Zone 1 fog-dominated microclimate. The study demonstrated early on that traits often treated as taxonomically diagnostic, such as pruina density, spine form, and body orientation, function as coordinated environmental responses calibrated to local conditions.
Fog regime and atmospheric structure
Phytogeographic and satellite-based analyses of the Chilean coastal desert demonstrate strong, repeatable structuring of vegetation into fog-defined belts and interior zones. These patterns form discrete fog-oasis ecosystems rather than a continuous vegetation band (Rundel et al. 1991; Schulz et al. 2011; Moat et al. 2021).
Oxygen isotope reconstructions from archaeological shells in the Paposo–Taltal corridor indicate that Early Holocene nearshore temperatures were approximately 3 °C cooler than present (Flores et al., 2018). This pattern is consistent with changes in coastal upwelling structure and suggests cooler conditions potentially more favorable to fog formation during the period when many present-day Copiapoa populations likely established.
Long-term shifts in fog frequency and cloud structure are associated with vegetation decline across these corridors, including documented impacts on Copiapoa populations.
Fog frequency in the Atacama Desert is not constant. Long-term satellite analyses show that it varies across interannual and decadal timescales, with strong links to large-scale ocean–atmosphere dynamics such as ENSO. These shifts are uneven across the landscape. In some sectors, fog occurrence has increased near the coast while declining at higher elevations, likely reflecting changes in the thermal inversion layer.
Because fog represents the dominant moisture input in many Copiapoa systems, this variability creates ecological uncertainty for recruitment and long-term population stability. These habitats operate close to their functional limits of moisture availability. Even modest changes in moisture input, fog duration, or surface conditions can disproportionately affect whether seedlings establish.
Why new plants rarely establish
Seed establishment in Copiapoa is constrained not only by moisture availability but also by light. Experimental work across Cactaceae shows that small-seeded taxa exhibit strong positive photoblastism, requiring light exposure for germination (Flores et al. 2011; Barrios et al. 2020).
This ties recruitment success to open surface microhabitats where fog and dew pulses coincide with high irradiance. Burial by shifting sand, caliche crusting, or surface disturbance can suppress regeneration even when adult plants persist, reinforcing the fragility of fog-oasis recruitment systems.
Across Cactaceae, optimal germination for most species falls between 20 and 30 °C / 68 and 86 °F (Barrios et al. 2020). However, at least one Copiapoa taxon, cinerea var. haseltoniana (now placed as Copiapoa gigantea f. haseltoniana following Larridon et al., 2015), shows peak germination at around 15 °C / 59 °F (Seal et al. 2017), consistent with adaptation to the cool, fog-buffered conditions of the coastal Atacama. Species adapted to cooler germination windows may be particularly vulnerable to warming, not because they are near a thermal ceiling, but because rising temperatures shift conditions away from the narrow optimum their seeds require. This risk is likely greatest on inland and north-facing exposures, where fog buffering is weakest and thermal loading is highest.
Direct field evidence from Copiapoa shows that population structure is not controlled primarily by competition, but by survival. Field studies of Copiapoa cinerea f. columna-alba populations at Pan de Azúcar found no evidence that plants compete strongly with one another for water or space (Gulmon et al. 1979). Instead, population density appears to be limited by the ability of plants to survive long dry periods, which can last for several years.
Water storage increases quickly with plant size. A stem about 22 cm / 8.7 in tall can survive for roughly 143 days without new moisture, while a small 2 cm / 0.8 in plant can survive only about 48 days under the same conditions. Smaller plants simply do not have enough stored water to survive extended dry intervals.
Plants below about 1.5–2 cm / 0.6–0.8 in in diameter were not found in any of the studied populations, even after careful searching. This suggests a critical survival threshold below which plants are unlikely to persist between moisture events.
The original study interpreted these dry intervals as gaps between rainfall events, because rainfall was assumed at the time to be the main water source. Current understanding shows that fog provides a major portion of moisture in many Copiapoa habitats. Periods of failed establishment are therefore better understood as intervals when both rainfall and fog input are insufficient.
Root studies show that Copiapoa has a shallow, spreading root system, with no roots deeper than about 8 cm / 3.1 in. Roots extend laterally across the soil surface, often covering an area comparable to or greater than the plant’s footprint. Root systems frequently overlap between individuals, further supporting that population density is not limited by competition. This architecture supports rapid capture of surface moisture rather than access to deeper water reserves (Gulmon et al. 1979; nomenclature reflects original usage).
In addition to these physiological constraints, rare large-scale disturbance events can reset entire populations. Geo-archaeological evidence indicates that the Atacama Desert coast has experienced rare, high-magnitude tsunami events, locally exceeding known historical tsunamis, capable of transporting marine sediments inland and reworking coastal substrates (León et al. 2019). In a system where Copiapoa populations are spatially restricted and slow to re-establish, such events represent a plausible mechanism for localized extirpation. This may help explain the absence of populations in otherwise suitable fog-supported habitats, where environmental conditions remain favorable but historical disturbance has reset the ecological system.
Together, these findings show that Copiapoa populations are shaped primarily by survival under environmental constraint. Establishment is the central bottleneck, and small shifts in moisture, temperature, or disturbance can determine whether recruitment occurs at all.
Longevity and recruitment failure
In fog-dominated systems, persistence depends on stable fog and condensation regimes rather than total precipitation. Small shifts in fog frequency, thermal load, or the timing and duration of condensation events can disrupt recruitment, moving populations from long-term persistence to demographic failure across generational timescales (Jordan & Nobel 1981; Thompson et al. 2003).
Rainfall input on the same coast is similarly episodic. The composite precipitation record assembled for Pan de Azúcar from regional stations and an in-park pluviometer shows a long-term mean near 13 mm per year together with extended gaps in which significant rain failed to arrive at all, including a roughly four-decade window from the mid-1940s to the mid-1980s with few measurable events (Thompson et al. 2003). Where fog and rainfall both supply moisture, the failure of either input for a sufficient period can produce a recruitment gap that becomes visible in the adult population only decades later .
In some coastal areas, large adult Copiapoa continue to survive even when conditions may no longer support reliable seedling establishment. These populations can be thought of as grandfathered stands: older plants remain alive even though new plants can no longer establish reliably under current conditions.
Mature plants have larger water reserves and are better able to tolerate heat and drought. Seedlings and young plants do not have this advantage and are much more likely to die during long dry periods.
A population made up mostly of older plants can therefore look stable while producing few or no new individuals. Over time, this leads to slow decline, where survival depends on existing plants rather than new growth.
🔴 Field Note: The presence of large, aging plants without younger individuals may indicate a population under stress. These stands can represent systems in gradual decline, where current conditions no longer support regeneration even though older plants remain alive.
Soil biology as a possible establishment filter
In addition to atmospheric and thermal limits, soil conditions may also affect whether seedlings can survive in the Atacama Desert.
Studies of hyper-arid soils in northern Chile show extremely low microbial life, limited diversity, and very few fungal partners. In other desert systems, even highly stress-tolerant plants struggle to establish when these microbial partners are missing.
For Copiapoa, direct data on soil microbiomes are still limited. However, existing findings suggest that soil biology may play a role in seedling survival by helping manage salt stress, water balance, nutrient availability, and root function at the soil surface. In areas where microbial communities are sparse or absent, this may add another barrier to successful establishment.
This remains a working hypothesis and has not yet been directly confirmed in Copiapoa.
Aguadas: Groundwater Wetlands of the Coastal Desert
The emphasis on fog throughout this account of Copiapoa habitat should not leave the impression that the coastal Atacama Desert is entirely without liquid water.
Scattered through the coastal quebradas and interior drainages are aguadas, points where subterranean water surfaces as fresh-to-brackish pools and seeps. These are groundwater emergence points, sites where geological structure forces water to the surface: more permeable units juxtaposing less permeable bedrock or finer-grained sedimentary units push accumulated groundwater upward until it breaks out (Sáez et al. 2016). The same basic mechanism operates from the inland Domeyko Range, where it has been documented in detail with radiometric chronology and sedimentological cross-sections, to the coastal Cordillera de la Costa, where quebrada napas surface at named aguada points within the drainage system.
Functioning as the Atacama's equivalent of desert oases, aguadas have concentrated biodiversity, human settlement, and travel routes along this coast for millennia, though they are far smaller and more hydrologically fragile than the deep-aquifer oases of other desert systems.
Copiapoa and the aguadas: neighbors, not dependents
It would be natural to assume that Copiapoa growing near these water points draw moisture from them. They do not. The two communities coincide spatially because both are expressions of the same underlying condition, a fog-favored quebrada system, rather than because one feeds the other. The Camanchaca sustains the Copiapoa communities on the fog-influenced slopes; the aguadas sustain a fundamentally different plant assemblage in the low quebrada floors where groundwater rather than fog is the dominant ecological driver.
The aguada vegetation is obligately tied to permanent water and occupies the moist channel floor and its margins. Copiapoa occupy the dry rocky slopes, terraces, and interfluves above and around these features, obtaining their moisture atmospherically from the Camanchaca, not from the substrate. Multi-year instrument data collected across five meteorological stations in Parque Nacional Pan de Azúcar confirm that the park may go without measurable rainfall for as long as a decade at a time (Thompson et al. 2003). Copiapoa survive those rainless decades because they draw on fog, not on rainfall and not on the groundwater that feeds the aguadas. A Copiapoa rooted in the permanently damp ground of an aguada margin would not thrive but rot; the entire physiological strategy of the genus is built around capturing transient atmospheric moisture and then drying completely.
So the Copiapoa and the aguadas are best understood as neighbors drawn to the same favorable setting for entirely different reasons. The geomorphology that funnels Camanchaca into a quebrada also creates the catchment within which infiltrated water can accumulate and resurface. Both the wetland at the valley floor and the cacti on the surrounding slopes are consequences of a fog-favored landscape, not of each other.
Pan de Azúcar: evidence from the field
The distinction is well-illustrated by the ecology of Parque Nacional Pan de Azúcar, where both communities have been studied in detail. Within the park, interior plant life away from the fog zone is limited to sites of aguada occurrence or to areas of concentrated runoff in arroyos (Thompson et al. 2003). Fog influence along this coast is highly locality-dependent: quebrada geometry, coastal exposure, and proximity to the marine inversion layer determine how far fog penetrates and at what elevations it condenses. At sites like Paposo and Pan de Azúcar, Camanchaca influence can occur essentially at sea level where coastal relief funnels and concentrates the inversion layer. What holds across localities is the ecological distinction, not a fixed elevation threshold: the aguada communities are sustained by groundwater, and Copiapoa communities by atmospheric fog moisture, regardless of the elevation at which that fog arrives.
The park contains ten named aguadas, covering roughly 84,200 square meters in total, roughly 0.02 percent of the park's area, classified as standing-water, watercourse, or mixed types (CONAF, park documentation). Despite their tiny footprint they support roughly 8 percent of the vascular plant species recorded in the park (Rundel et al. 1996), including obligate-water plants with permanent moisture requirements: salt-tolerant grasses such as Distichlis spicata, reeds including Juncus acutus, halophytes including Sarcocornia fruticosa and Frankenia chilensis, and the Atacama toad (Rhinella atacamensis), whose breeding population requires reliable year-round water rather than ephemeral atmospheric moisture. None of these species appear in the fog-fed lomas or Copiapoa slope communities. They are ecologically and hydrologically separate assemblages sharing a fog-favored landscape without sharing a water source.
Distribution and deep history
The aguadas are not a unique feature of Pan de Azúcar. They are a recurring element of the Atacama coastal desert wherever geological and topographic conditions permit groundwater to surface. The Taltal district alone contains at least 40 documented coastal spring points (Contreras et al. 2007, cited in Mendez et al. 2020), and the broader Domeyko Range contains at least five major groundwater discharge systems with stratigraphic records extending back 15,000 years (Sáez et al. 2016). What distinguishes Pan de Azúcar is not the existence of aguadas but the degree of documentation, protection, and scientific study they have received.
The aguadas have structured human occupation of this coast for at least 12,000 years. Archaeological records of the Taltal district indicate that the first coastal settlements were always associated with these freshwater spring points, and the aguadas served as replenishment spaces and social nodes for caravanning communities linking the coast to the Andean interior through the Early Holocene and beyond (Mendez et al. 2020; Núñez and Varela 1967). That 12,000-year continuity makes the aguadas one of the most durable organizing features of human life on this coast, more persistent than any particular settlement, mining enterprise, or colonial boundary.
The groundwater that feeds these coastal aguadas is itself largely ancient. Isotopic and radiocarbon analysis of small springs on the western face of the Cordillera de la Costa near Antofagasta indicates that the water discharging today reflects a period of significantly greater recharge between roughly 5,000 and 3,000 years ago, likely driven by more frequent and intense ENSO events bringing warm Pacific currents southward along the Chilean coast (Herrera and Custodio 2014). Current rainfall in the coastal range averages less than 1 mm per year in many locations and fog condensation, while critical to vegetation, does not contribute meaningfully to aquifer recharge (Herrera and Custodio 2014). The aguadas are in this sense drawing down inherited water, and the long-term decline in spring discharge documented in historical and pre-historical records is consistent with this interpretation.
Conservation significance
The aguadas are among the most disturbance-sensitive ecosystems in the park, vulnerable to contamination, vegetation destruction, and changes in water flow caused by groundwater extraction outside the park's boundaries, from which recovery is slow and uncertain (Peña-Cortés et al. 2006; CEA 2011). Because their recharge originates upstream and frequently outside the protected area, the aguadas illustrate a principle that runs throughout Copiapoa habitat conservation: the pressures that matter most often arrive from outside the boundary that is meant to offer protection.
The threat is not hypothetical and not new. In the Taltal district, the aguadas were progressively piped, diverted, and legally contested throughout the nitrate mining boom between 1885 and 1920, with pipeline infrastructure extending from the Aguada de Varas to the Mineral del Guanaco by 1885 and all known aguadas under modern extraction infrastructure by 1920 (Mendez et al. 2020). Further inland in the Domeyko Range, recent mining trenches have cut directly into Quaternary groundwater discharge deposits to extract water, exposing and disrupting sedimentary records of spring activity stretching back 15,000 years (Sáez et al. 2016). The sedimentary record itself shows a trend toward decreasing groundwater discharge over the last few centuries, suggesting these systems are already under climate pressure independent of any extraction (Sáez et al. 2016).
The aguadas that make the Pan de Azúcar landscape biologically extraordinary are not resilient features. They are finite, geologically controlled, externally recharged, and historically contested. Their continued presence in the landscape is not guaranteed by the park boundary that surrounds them.
Source Basis: Fog-structured vegetation corridors and population persistence draw on Rundel et al. (1991), Schulz et al. (2011), Moat et al. (2021), and Gulmon et al. (1979). Germination constraints follow Flores et al. (2011) and Barrios et al. (2020). Longevity and recruitment failure follow Jordan and Nobel (1981) and Thompson et al. (2003). Soil biology is treated as a working hypothesis.
Aguada hydrology follows Sáez et al. (2016). Ecological distinction between aguada and Copiapoa fog-belt communities, and Pan de Azúcar climate data, follow Thompson et al. (2003). The aguada flora inventory and 8 percent vascular plant species figure are attributed to Rundel et al. (1996) via park documentation; confirm against the primary paper before treating as a hard number. Historical water use and mining extraction follow Mendez et al. (2020). Conservation vulnerability cites Peña-Cortés et al. (2006) and CEA (2011) via park documentation; both remain secondary citations.
Full citations are on the Reference page.
Habitat interpretation guide
Copiapoa do not exist in isolation. The surrounding biological community and geological setting often provide the clearest clues to fog influence, ecotype, and long-term habitat conditions.
In coastal and lower transitional zones, persistent lichenized fungi, cyanobacterial films, cryptogamic crusts, and other biological surface communities usually indicate stable fog input. At anchors such as Paposo, Blanco Encalada, Pan de Azúcar, and the El Soldado Corridor, these signals often correspond to Zone 1–2 fog regimes.
Vascular fog indicators can strengthen this interpretation. Within the Taltal corridor, species such as Tillandsia landbeckii and Oxalis gigantea indicate reliable atmospheric moisture. By contrast, stress or dieback in columnar cacti such as Eulychnia iquiquensis, especially on exposed coastal escarpments near Caleta Cifuncho, may indicate declining fog reliability and increasing thermal or radiative stress.
Biological Inversion Line
One of the most useful field markers is the abrupt disappearance of lichenized fungi from exposed rock surfaces. Near Cerro Perales and Quebrada Botija, this collapse marks the upper margin of biologically usable fog input. This guide refers to that threshold as the Biological Inversion Line. Above it, Copiapoa survival shifts away from sustained fog interception and toward extreme moisture limitation, mineral-mediated condensation, micro-scale lithic moisture retention, and specialized root strategies.
Substrate can be just as important as fog. In the El Soldado Corridor, high-albedo granitic surfaces exclude much surrounding vegetation while favoring specialized Copiapoa forms with heavy white epicuticular wax. In the Chañaral–Copiapó Gateway, extensive caliche crusts restrict rooting to fractures and margins rather than the crust itself.
These habitat signals should be read together. Fog indicators, slope, aspect, substrate, and surrounding vegetation often reveal the ecological setting more reliably than elevation or surface appearance alone.
Source Basis: Fog-oasis structuring and vegetation belts follow Rundel et al. (1991) and Moat et al. (2021). Microbial structuring within the Quebrada Botija corridor follows Cayo et al. (2025). Full citations are on the Reference page.
Flooding and substrate stability shape Copiapoa distribution
Geographic Anchors and Fog Corridors
Geographic framework
To organize the distribution of Copiapoa across the Atacama Desert coastal range, this guide uses a three-level geographic framework:
- Fog Oasis System: the broader Atacama Desert coastal fog belt extending from southern Peru to north-central Chile
- Geographic Anchors: regional environmental corridors representing recurring fog regimes, lithologic transitions, and elevation structures
- Locality Populations: individual sites, field-numbered populations, or collector localities within each anchor region
This structure reflects how Copiapoa populations occur in nature: not as isolated points, but as components of larger fog-structured landscape systems.
Why anchors are used
Attempting to catalog every micro-locality across the Atacama Desert coast can obscure the broader ecological structure that governs Copiapoa distribution. The anchor system highlights recurring environmental corridors that shape population structure across the genus.
At this scale, Copiapoa populations are best understood as components of larger landscape systems defined by fog interception patterns, coastal topography, lithologic substrate regimes, and elevation-controlled moisture gradients.
Anchors do not represent species boundaries and are not intended as exhaustive distribution maps. Each anchor encompasses multiple microhabitats and local populations that occupy a shared fog corridor or environmental gradient. Together, they bracket the major fog corridors, substrate transitions, and topographic discontinuities that structure both morphological variation and documented phylogeographic patterning within the genus.
The anchor framework therefore provides a stable geographic reference structure for interpreting locality records, phenotype variation, and ecological differentiation.
Fog oasis context
The broader fog-oasis framework underlying these anchors is informed by the remote-sensing analysis of Moat et al. (2021), who used a 20-year archive of MODIS satellite imagery to map desert fog oasis ecosystems along the Pacific coastal belt of Peru and Chile.
Their study identified more than 17,000 km² of fog-dependent vegetation distributed across more than 900 discrete fog oasis patches. These ecosystems occur as fragmented ecological islands shaped by elevation, slope, distance from the coast, atmospheric moisture interception, and the geometry of the coastal escarpment itself.
Along much of the Atacama Desert margin, the coastal cliff and Coastal Cordillera restrict farther inland movement of the marine stratocumulus layer. Where the escarpment is interrupted by quebradas or lower-relief corridors, fog can penetrate inland more effectively. GOES-based studies identified these corridors directly, and a later satellite-based climatology using machine learning techniques is consistent with their recurrence as landscape-scale features, with enhanced fog occurrence relative to adjacent unbroken escarpment sections. The strongest penetrations occur during austral winter advective events (Farías et al. 2005; Böhm et al. 2021).
The geographic anchors used here therefore reflect not only fog frequency at a given elevation, but also the topographic structure that determines where fog can reliably reach the land surface.
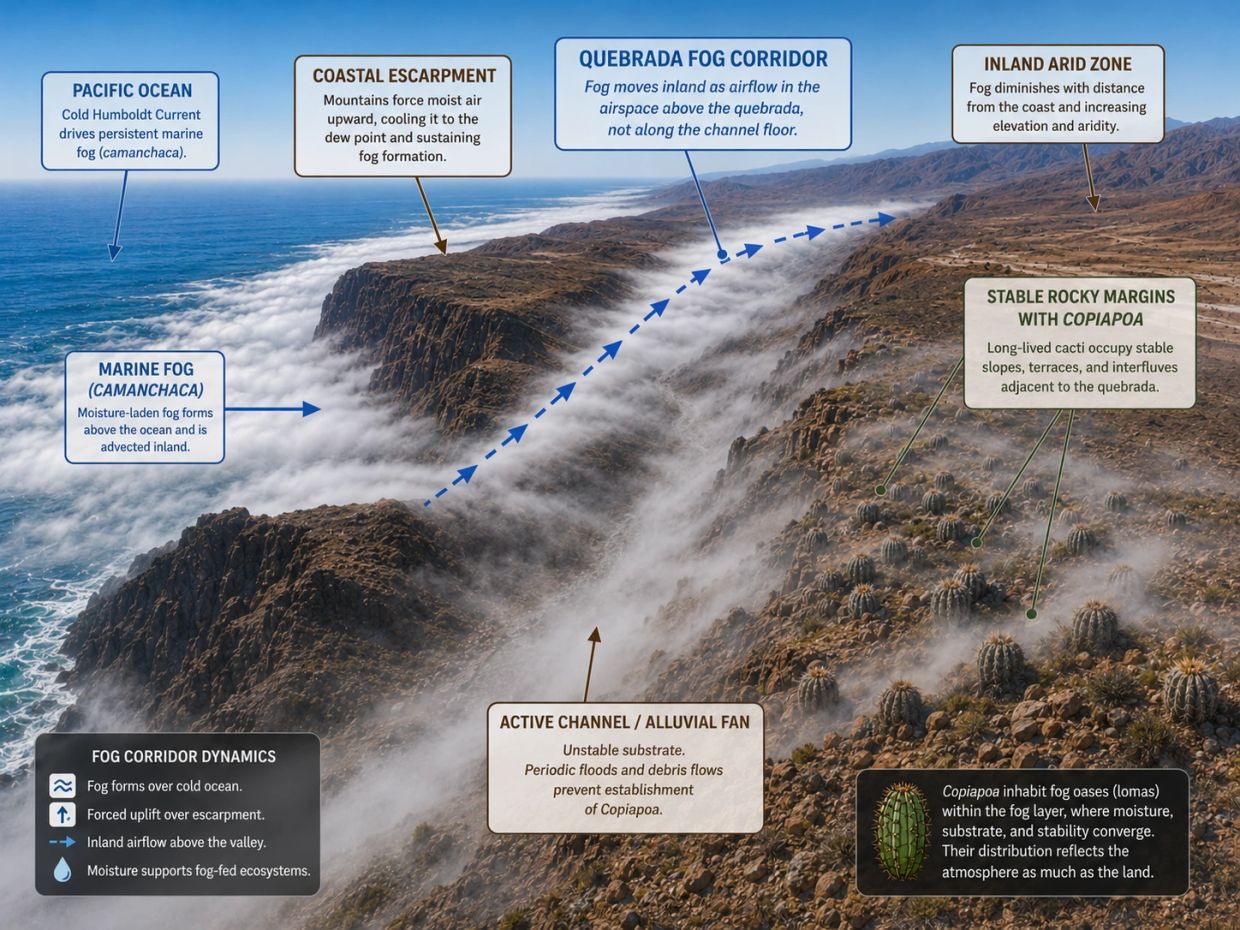
Quebradas as fog corridors and ecological filters
Along the Atacama Desert coast, the Cordillera de la Costa rises steeply from the Pacific, intercepting the marine fog layer and limiting inland moisture movement. Where quebradas cut through this barrier, they create topographic corridors that allow Camanchaca to penetrate inland along narrow drainage profiles.
These corridors concentrate atmospheric moisture and help support fog-dependent vegetation, including Copiapoa, in terrain that would otherwise be too arid for establishment.
However, Copiapoa do not usually occupy active quebrada channels. Periodic debris flows, sediment movement, and mechanical disturbance make active channels unsuitable for plants whose individuals may persist for many decades to centuries. Instead, populations establish on stable rocky margins, abandoned terraces, cemented interfluves, and adjacent slopes where fog input coincides with long-term substrate stability.
This distinction is central: the quebrada is the fog pathway, but the plant habitat is the stable margin beside it. A drainage may channel abundant fog, but if its active floor is repeatedly reworked, Copiapoa cannot persist there. Dense populations develop only where reliable atmospheric moisture overlaps with geomorphically stable rooting surfaces.
This framework explains why Copiapoa often flank quebradas rather than occupy them directly. Their shallow, laterally spreading roots are adapted to rapid capture of surface moisture from fog, dew, and brief wetting events, but they require stable surfaces long enough for seedlings to reach a survival threshold. Field studies at Pan de Azúcar found that very small plants below approximately 1.5–2 cm were absent from surveyed populations, suggesting that early establishment is the main bottleneck.
The same corridor-margin pattern appears across much of the genus range, although the dominant moisture regime changes by latitude. In the northern hyper-arid core, quebradas such as Botija, Izcuña, El Médano, and San Ramón function primarily as fog conduits. Farther south, at sites such as Llanos de Challe and Huasco, quebradas still structure Copiapoa habitat, but within a fog-rainfall hybrid regime.
Near Tocopilla, populations thin out, likely reflecting reduced fog reliability among other contributing factors. Near Coquimbo, quebradas remain functional landscapes, but the dominant moisture regime shifts toward semi-arid rainfall rather than fog.
Together, these northern and southern limits bracket the functional fog-corridor system that structures most Copiapoa distribution. The quebrada model is therefore not simply a list of drainage names, but a way to understand how atmospheric moisture, topography, geomorphic stability, and plant longevity interact across the Atacama Desert.
Principal geographic anchors
The following anchors outline the primary environmental corridors that structure Copiapoa distribution along the Atacama Desert coast.
The quebradas listed under each anchor are not presented as simple habitat sites, but as landscape structures: fog conduits, drainage corridors, historical travel routes, and geomorphic filters.
Copiapoa populations generally occupy stable rocky margins, terraces, and interfluves associated with these systems rather than active channels.
- Tocopilla (Northern Perimeter)
- Located within the major Atacama Desert arid divide identified by Moat et al. (2021), a sector extending roughly from the Peru–Chile border to Antofagasta where verdant fog oases become sparse and fragmented, persisting only as small, isolated patches.
- Contributing Quebradas: Quebrada Barriles, Quebrada Mamilla. These drainages define the coastal escarpment structure at the northern distributional edge but Copiapoa tocopillana has not been reliably relocated in recent field surveys despite multiple attempts (Hoxey 2001, 2015). This anchor functions as a distributional boundary reference rather than a documented fog-corridor habitat.
- Quebrada Botija (High Montane Specialist)
- Within documented fog oasis terrain in the Paposo–Taltal corridor. Not individually resolved at the regional scale of Moat et al. (2021).
- Contributing Quebradas: Quebrada Botija (self-naming anchor). Copiapoa decorticans, C. ahremephiana, and C. solaris are documented within this drainage. El Medano style rock art confirms its use as a coast-to-interior corridor since at least the mid-Holocene (Monroy et al., FONDECYT 1151203).
- Blanco Encalada (Coastal Solaris Counterpoint)
- Within documented fog oasis terrain along the northern Chilean coastal belt. Not individually resolved in the regional MODIS mapping.
- Contributing Quebradas: Quebrada Izcuna. Copiapoa humilis subsp. variispinata grows in fragmented populations from the coast to 4.5-6.5 km inland along this quebrada, with subpopulations at higher altitudes receiving direct fog input (Ritter; llifle FK 382; Monroy et al., FONDECYT 1151203).
- Paposo (Northern Coastal Biodiversity Peak)
- Corresponds directly to the largest contiguous fog oasis complex documented in Chile by Moat et al. (2021), exceeding 610 km² and representing the highest concentration of fog oasis vegetation along the Chilean Atacama Desert coast.
- Contributing Quebradas: Quebrada El Medano, Quebrada de la Plata, Quebrada Paposo, Quebrada Matancilla, Quebrada Bandurrias. These drainages cut through the Cordillera de la Costa within this fog oasis complex. Aguadas within them support vegetation and historically supported livestock, confirming their role as fog conduits with permanent or seasonal water availability (Monroy et al., FONDECYT 1151203; Molina 2007a; Vidal Gormaz 1881).
- Taltal (Cinerea Type Locality)
- Located within the Paposo–Cifuncho fog oasis corridor, the most extensive contiguous fog oasis system documented in northern Chile by Moat et al. (2021). The Taltal region forms part of this larger coastal fog belt and represents the historical type locality for Copiapoa cinerea.
- Contributing Quebradas: Quebrada de Taltal, Quebrada Cascabeles, Quebrada San Ramon, Quebrada Portezuelo, Quebrada at Caleta Tortolas. Columna-alba populations occupy these quebradas running toward the Pacific, establishing within the drainage corridors but stopping short of the immediate beach zone (Monroy et al., FONDECYT 1151203; Molina 2007a; Hoxey 2004).
- Cerro Perales (High Montane Fog-Shadow Transition)
- Within the Paposo–Taltal fog corridor. Represents a transitional high-elevation environment where fog influence begins to weaken inland.
- Contributing Quebradas: Cerro Perales is positioned on the upper margin of the Quebrada Taltal drainage system. The summit overlooks Quebrada Taltal (Hoxey 2004), and the finest stands of white-waxed Copiapoa cinerea occur on the slope between the summit and the quebrada floor. The Perales locality is documented as a named aguada in the coastal mountains (Vidal Gormaz 1881).
- Esmeralda (Volcanic Heat-Battery Substrate)
- Within documented fog oasis terrain south of the Paposo complex. Not individually resolved at the regional mapping scale.
- Contributing Quebradas: Quebrada Guanillos (also rendered Huanillos on local signage), Quebrada Tigrillo. Both drain through the Esmeralda sector between Las Lomitas and the coast. Guanillos contains the full characteristic assemblage: C. longistaminea, C. grandiflora, C. esmeraldana, and C. laui (Hoxey 2003, 2010; KK 1385).
- Pan de Azúcar (Granitic Coastal)
- A major fog oasis biodiversity center with 293 recorded plant species in published surveys, representing the second highest documented diversity for any Chilean fog oasis region.
- Contributing Quebradas: Quebrada del Castillo, a named drainage within the national park where Copiapoa populations are documented along the trail system accessible from Camino C-110 (CONAF; Hoxey 2010).
- El Soldado Corridor (High-Albedo Mirror Substrate)
- Within documented fog oasis terrain between Pan de Azúcar and Chañaral. Not individually resolved at the regional MODIS scale.
- Contributing Quebradas: Quebrada La Madera (between Pan de Azucar and the Guanillos/Esmeralda sector). South toward Chanaral, the corridor is structured by short coastal drainages between Punta Animas, Los Toyos, Puerto Flamenco, Rada Blanca, and Punta Zentena. Unlike the Paposo or Taltal corridors, this sector is characterized by minor drainages across high-albedo, Miocene-stable interfluves rather than a dominant fog-channeling quebrada (Hoxey 2003, 2010).
- Chañaral–Copiapó Gateway (Phylogeographic Transition Zone)
- This corridor marks a major environmental transition between the northern Atacama Desert fog belt and the southern coastal desert systems. While fog oases persist farther south, the Chañaral–Copiapó region represents an important ecological and phylogeographic inflection point within Copiapoa, where fog influence weakens and the structure of the coastal desert begins to change.
- Contributing Quebradas: This anchor operates on different logic from the fog-corridor anchors further north. The Rio Copiapo is a broad fluvial valley functioning as a biogeographic filter, and the named quebradas here are Cordillera de Domeyko interior drainages: Quebrada La Encantada, Quebrada Dona Ines, Quebrada Patos Cerrados (now Jardin), Quebrada Chanaral Alto, Quebrada Paipote (Molina 2007a; San Roman 1896; Philippi 1860).
- Llanos de Challe (Southern Refuge)
- Classified as a transitional fog oasis system where fog-dependent desert vegetation grades into Mediterranean-type sclerophyllous communities at the southern ecological boundary of the Atacama Desert fog oasis system.
- Contributing Quebradas: Quebrada Mala, Quebrada Barracota. Within the national park, C. dealbata, C. echinoides, and Eulychnia breviflora are documented on low hills accessible from the ranger station (Hoxey 2010, 2013, 2019).
- Huasco / Carrizal Bajo (Southern Perimeter Reference)
- Corresponds to the southern ecological transition of the fog oasis system identified by Moat et al. (2021), marking the terminal extent of the primary Atacama Desert fog-dependent vegetation corridor.
- Contributing Quebradas: Quebrada Cruz Grande, Quebrada Honda, Quebrada de Los Hornos, Quebrada Juan Soldado (Aguilera 2014). Copiapoa fiedleriana is documented near Huasco (Hoxey 2013).
- Coquimbo (Southern Transition Reference)
- Located south of the primary Atacama Desert fog oasis system and outside the geographic scope of the Moat et al. (2021) mapping analysis.
- Contributing Quebradas: Quebrada Santa Gracia, Quebrada Marquesa (Soto et al. 2017). These drainages mark the southern transition where the fog-dominated moisture regime gives way to semi-arid rainfall. Like Tocopilla at the northern boundary, Coquimbo defines the genus range limit not by the absence of quebradas but by the transition of the moisture regime that gives them ecological meaning.
The Copiapó transition
The Chañaral–Copiapó sector corresponds to one of the most significant north–south transitions documented in the genus. Molecular phylogenetic analyses identify a major plastid clade division broadly corresponding to this sector, separating predominantly northern and southern evolutionary lineages.
This transition also coincides with a bioclimatic shift. North of Copiapó, conditions become increasingly hyper-arid, with stronger reliance on persistent marine fog and reduced inland moisture buffering. Southward, precipitation regime, vegetation structure, and substrate mosaics begin to shift.
Geomorphically, the Copiapó Valley forms a broad discontinuity within the coastal range. In its coastal sector, the valley widens substantially and is dominated by unstable sandy substrates. These conditions likely reduce the effectiveness of the short-distance, ant-mediated seed dispersal typical of Copiapoa. Periodic flood events further limit long-term plant establishment across the valley floor.
The valley is not an absolute barrier. It functions as a soft biogeographic filter and coincides with one of the clearest phylogeographic inflection points within the genus.
For this reason, the Chañaral–Copiapó Gateway is treated here as a primary structural anchor in both ecological and evolutionary context.
Chañaral Conservation Concern
One additional variable in this corridor remains underexamined. Chañaral Bay carries one of the world’s most extensively documented marine mine-tailings contamination events, with approximately 150 million tons of copper mine tailings discharged directly into the bay during El Salvador mine operations from 1938 to 1990. Persistent sediment contamination continued after flood-resurgence events in 2015 and 2017.
Hydrochemical analyses of Camanchaca in the Chañaral sector demonstrate elevated concentrations of dissolved metals, including copper and arsenic, derived from coastal contamination sources (Bonnail et al. 2018). Because this fog is advected inland across the same coastal slopes that host Copiapoa populations, atmospheric deposition of dissolved metals via fog and associated aerosols represents a plausible but untested exposure pathway.
These findings indicate that Atacama Desert coastal fog systems are not uniformly chemically pristine. Localized anthropogenic contamination may create additional environmental pressure within specific coastal corridors.
Because long-term population stability in Copiapoa depends on successful recruitment rather than adult survival alone, even low-level contamination in fog-dependent systems could influence germination and early establishment without being immediately visible in standing adult populations. This mechanism is explored in greater detail in the Coastal Contamination and Fog-Mediated Transport section.
Floristic analysis and connectivity
Published floristic similarity analyses of the coastal Atacama Desert indicate strong affinity between Paposo and Taltal, supporting their interpretation as components of a coherent northern coastal fog corridor rather than isolated fog systems (Larraín Barrios 2007; Muñoz-Schick et al. 2001).
These analyses, based on Jaccard similarity indices, show greater floristic overlap between Paposo and Taltal than between Paposo and more southern fog systems. This reflects shared fog structure, elevation bands, and exposure regimes.
Paposo is further distinguished by a high proportion of locally restricted taxa, supporting its designation as a regional biodiversity anchor despite the presence of endangered subpopulations.
In contrast, anchors such as Blanco Encalada and Quebrada Botija function as diagnostic bookends. They illustrate how a single lineage such as Copiapoa solaris shifts morphology, physiology, and rhizosphere ecology as it transitions from coastal Zone 1–2 fog regimes into high montane Zone 4 thresholds.
Source Basis: Fog corridor structure and topographic channeling follow Farías et al. (2005), Böhm et al. (2021), and Moat et al. (2021). Archaeological and historical corridor use draws on Monroy et al. (FONDECYT 1151203) and Molina (2007a). Survival threshold and root architecture follow Gulmon et al. (1979). Floristic zonation follows Larraín Barrios (2007) and Muñoz-Schick et al. (2001). Phylogeographic context follows Larridon et al. (2015). The inference that mechanical recurrence in active channels excludes long-lived plants has not been experimentally tested for Copiapoa specifically. Full citations are on the Reference page.
The paradox of flooding in the hyperarid Atacama Desert
The flooding that keeps Copiapoa from growing on active quebrada floors can seem paradoxical given how little rain reaches the Atacama coast. The answer lies upstream.
In the large quebradas that descend from the Andes to the Atacama coast, the flood water that scours the channel often originates far inland. Rainfall in the precordillera and at mid elevations, frequently during ENSO-linked or cutoff-low events, generates runoff that funnels down the drainage network and arrives near the coast hours later, sometimes under clear skies. Much like a flash flood in the Arizona desert, where a wash can run high and fast under a clear sky because the rain fell on distant mountains, a person standing at one of these quebrada mouths may never see the weather that produced the flood.
Smaller catchments in the Coastal Cordillera behave differently in this one respect. On the rare occasions they do run, their flows are fed by storms striking the coastal range directly, so a person standing in the drainage is usually experiencing the same weather that produces the runoff. The channel response is much the same either way, brief and sediment-charged, but the source of the water is not.
Why these floods are unusual - and powerful
These events differ from a typical Arizona wash in two ways that matter for plant habitat.
First, they are far rarer. The pattern appears to consist of moderate floods every few decades, punctuated by much larger events that may not recur for a century or longer. Long dry intervals allow loose sediment to accumulate in channels and valley fills. When a flood finally arrives, it does not simply move water downslope. It remobilizes years or decades of stored material, eroding its own bed and banks and sometimes widening the active channel substantially in a single event.
Second, many of the larger floods take the form of debris flows or highly sediment-laden runoff rather than simple muddy water. These dense mixtures of mud, sand, and rock carry enormous geomorphic force. The flows are also brief and intense. With little soil storage, almost no vegetation to slow runoff, and no sustaining baseflow, much of the water and channel reworking can pass in a matter of hours before the drainage returns to dry.
How the landscape builds over time
Each rare event delivers a large pulse of sediment that is deposited as the flow loses gradient and spreads near the coast. Over time, these deposits build the alluvial fans found at the mouths of many quebradas.
Importantly, the size of these fans is not evidence of frequent flow. They are the cumulative product of many rare, sediment-charged events spaced far apart in time. Between floods, material simply waits in the drainage system until the next major runoff event arrives.
In catchments affected by mining, that material is not always clean. Floods and debris flows can entrain mine tailings and metal-enriched substrate alongside natural sediment. Much of this material never reaches the ocean, instead being stored on alluvial fans and lower piedmont surfaces, where it may remain for decades before being redistributed by wind or future floods.
Why plants avoid the channels
Recovery from one of these events is slow. A major flood does not simply leave the channel dry again; it leaves it geomorphically disturbed. The time required for the channel to stabilize is long relative to the time a seedling needs to establish.
For long-lived desert plants such as Copiapoa, establishment is measured in decades rather than seasons. Even infrequent episodes of scouring, burial, and channel reworking are enough to repeatedly eliminate young plants before they become established. As a result, active channels and their lowest walls remain largely uncolonized.
The stable margins above the flood zone tell a different story. Terraces, slopes, and interfluves persist long enough for seedlings to survive the vulnerable establishment phase, which is why Copiapoa populations typically line the edges of quebradas rather than grow within them.
Ecotype Zone Framework and Evidence
For more than half a century, researchers, field explorers, and growers have documented strong links between Copiapoa morphology, population structure, and habitat conditions across the Atacama Desert. These observations established early recognition that much of the visible variation within the genus reflects environmental context as much as, and often more than, formal species boundaries.
Building on this foundation, copiapoa.com formalizes these long-standing observations into a unified four-zone ecotype framework. The model integrates verified locality data with modern molecular studies, satellite-derived fog climatology, and contemporary desert ecology. Rather than replacing earlier taxonomic or horticultural interpretations, the framework synthesizes them into a coherent structure that allows growers, researchers, and conservationists to interpret Copiapoa diversity consistently and transparently.
The framework reflects documented fog frequency, fog-water yield, ultraviolet intensity, thermal regime, and the known physiological limits of Copiapoa. Ecotype boundaries are defined by persistent environmental structure rather than informal growth-form descriptors or unverified elevation estimates. Long-term fog monitoring confirms that fog-dependent ecosystems occur as distinct fog oases with predictable elevation thresholds, providing a defensible climatic basis for ecotype zonation.
Framework note: This ecotype framework is presented as a working ecological model, not a rigid classification system. It is expected to evolve as higher-resolution fog data, isotope analyses, and genetic studies become available.
Locality Data, Elevation Accuracy, and Fog-Bank Precision
Locality information, particularly elevation, is frequently approximate or inconsistently reported. Descriptors such as "mountain form" often reflect steep terrain or inland exposure rather than true altitude and should not be treated as reliable ecotype indicators. Accurate elevation requires direct measurement using an altimeter, GPS, or modern mapping software. Visual estimates without instruments are commonly inaccurate.
High-resolution fog-collector networks and satellite analyses document a persistent marine fog belt extending from sea level to approximately 800 to 1,000 meters along much of the coast. Moisture availability declines sharply above the marine inversion layer, with fog-water yield collapsing rapidly above roughly 1,000 to 1,100 meters depending on locality and season. These recurring atmospheric bands correspond directly with the ecotype zones described here and provide a defensible basis for placing the lower boundary of the high montane ecotype near this elevation.
The following section explains how water availability changes with elevation and why survival above the fog belt requires more than fog alone.
Hydrological Evidence Supporting Ecotype Zonation
Stable isotope analyses and hydrological studies of Atacama Desert cacti reveal a consistent gradient in water-source use across the landscape. Near the coast, plants rely primarily on fog. Further inland and at higher elevations, hydration shifts toward dew, rare precipitation, and other non-fog atmospheric inputs. Dependence on fog declines predictably with distance from the ocean, elevation, and exposure, marking the transition from coastal to inland water economies.
Independent non-biological studies demonstrate a sharp inland and elevational decline in biologically available surface moisture. These findings closely align with the observed transition from fog-dependent coastal ecosystems to arid inland corridors and independently support the environmental gradients used to define Copiapoa ecotype zones.
Beyond atmospheric sources, geochemical and microbiological research shows that hygroscopic minerals such as halite and gypsum can retain or generate liquid water internally through nano-pore capillary condensation, even when ambient humidity remains below the dew point. Endolithic microbial communities persist in hyper-arid Atacama Desert rock by exploiting this mineral-bound moisture, confirming that biologically accessible water can exist where fog and rain are absent.
Although direct mineral-mediated water uptake by Copiapoa has not been experimentally demonstrated, the persistence of high-elevation populations suggests that mineral-associated buffering pathways may function as part of a broader hydrological safety net under extreme moisture limitation. This mechanism plausibly explains the compact ribs, thick cuticles, and extremely slow growth characteristic of Zone 4 ecotypes.
Stable carbon isotope signatures from the Atacama Desert further confirm extreme water stress above the fog belt. These values rank among the more extreme signatures reported for terrestrial plants under chronic water stress, underscoring the severe hydric limitation experienced by inland and high montane ecotypes. Complementary soil studies show that organic material is uniformly scarce across the hyper-arid Atacama Desert and that rare biological inputs correlate with fog influence rather than rainfall or landform, further supporting the fog-structured ecological gradients used to define Copiapoa ecotype zones.
Taken together, these lines of evidence demonstrate that morphological divergence in Copiapoa reflects enduring hydrological and atmospheric structure rather than species boundaries, hybridization, or short-term climatic variation.
🔴 Scientific anchor: Spine pigmentation and architecture respond developmentally to sustained environmental conditions while remaining constrained by inherited genetic limits. Environment modulates expression. It does not redefine lineage without sustained population-level evolutionary change.
Quick Reference: Ecotype Zones
The four ecotype zones can be summarized before examining each in detail:
- Zone 1 Coastal Fog Belt
Persistent fog and stable atmospheric hydration.
Typical expression: white, wax-heavy bodies; broader, hydrated ribs; softer spination.
- Zone 2 Transitional Fog Belt
Intermittent fog with increasing dry intervals.
Typical expression: mixed coastal and inland traits; firmer epidermis; moderate spination.
- Zone 3 Inland Fog-Shadow
Low fog reliability with high radiative and thermal stress.
Typical expression: darker bodies; compact growth; stronger protective spination.
- Zone 4 High Montane
Fog-water collapse above the inversion layer.
Typical expression: ultra-slow growth; compact stress forms; reduced rib expansion.
These zones describe the plant’s baseline ecological strategy. Local substrate, slope, and mineral conditions can modify expression within any zone, but they do not change the underlying ecotype.
Source Basis: Fog frequency and elevation thresholds follow long-term collector networks and satellite climatology reviewed in Cereceda et al. (2008) and Moat et al. (2021). Hydrological gradients and water-source transitions draw on stable isotope analyses of Atacama Desert cacti. Mineral-bound moisture pathways follow endolithic microbial studies in hyper-arid systems. Full citations are on the Reference page.

Coastal fog is the lifeline of the Atacama Desert

Ecotype Zones Definitions
Modern fog research provides the foundation for defining Copiapoa ecotype boundaries. Long-term ground-based fog collectors, combined with satellite-derived cloud-frequency datasets, now allow the major environmental bands of the coastal Atacama Desert to be identified with far greater precision than was previously possible.
Ecotype boundaries are defined using persistent atmospheric structure together with repeatable, population-level morphological responses observed in wild populations. The four ecotype zones describe how Copiapoa morphology aligns with long-term patterns of fog immersion, humidity stability, solar exposure, and temperature regime across the Atacama Desert. While fog collectors measure droplet flux approximately 1 to 2 meters above ground, Copiapoa respond primarily to humidity, boundary-layer conditions, and fog immersion at the plant surface. Ecological interpretation therefore integrates measured fog trends with biological signatures expressed by plants over decades to centuries. Zone boundaries presented here follow combined biological and atmospheric indicators rather than fog-collector yield alone.
Ecotype zones define a plant's core architectural strategy, including water storage capacity, epidermal structure, rib dynamics, and growth form. Local substrate and terrain further modify expression within any zone, influencing color, rib emphasis, spine density, and surface reflectivity, but they do not change a plant's ecotype.
➤ Zone 1: Coastal Fog Forms (0 to 500 m)
Environmental context: This zone occupies the core of the coastal fog belt, where fog frequency is highest and fog-water yield peaks along many slopes. Fog immersion and condensation often peak between approximately 300 and 600 meters, depending on local topography, but strong fog influence extends down to near sea level in exposed coastal sectors.
Climatic data: Absolute humidity remains relatively high for the region, commonly falling in the range of approximately 7 to 12 g/kg along fog-influenced coastal sectors. These conditions persist several kilometers inland,
helping maintain relatively stable atmospheric hydration for coastal populations.
PAR: Heavily suppressed by fog. At Las Lomitas, Jung et al. (2020) recorded an annual mean PAR of 892.72 ± 727.53 µmol/m²/s and a maximum of 2,596 µmol/m²/s. Despite the Atacama's reputation for extreme solar exposure, measured PAR at this fog-dominated site falls within ranges commonly encountered in cultivation and many non-desert environments.
Traits:
- Heavy chalk-white epicuticular wax reflecting light and aiding fog interception
- Broad, water-rich ribs that expand readily with atmospheric hydration
- Soft, thin epidermis adapted to stable, humid conditions
- Compact silhouettes optimized for frequent fog exposure and low PAR
- Short, upright spines commonly associated with fog-exposed populations
Examples: Coastal forms of Copiapoa cinerea, dealbata, gigantea, marginata, fiedleriana, and many coastal humilis populations.
Primary anchors: Paposo, Blanco Encalada, El Soldado Corridor (coastal sectors), Pan de Azucar (coastal sectors), Chañaral–Copiapó Gateway (coastal sectors), Llanos de Challe (coastal sectors).
➤ Zone 2: Transitional Forms (400 to 800 m)
Environmental context: Fog persists within this band but with reduced density and shorter duration. Hydration occurs through intermittent fog pulses that are strongly dependent on slope exposure and airflow. Increased direct sunlight, stronger UV exposure, and wider diurnal temperature ranges progressively shape plants toward more stress-tolerant forms.
Climatic data: Absolute humidity remains within the coastal fog regime but exhibits greater temporal variability and more frequent dry intervals as inland air interacts with advective fog.
Traits:
- Moderate to thin epicuticular wax, often restricted to the apex or newest growth
- Firmer, more UV-tolerant epidermis with grey-green or green tones
- Shift from globose to short cylindrical forms in many populations
- Spination stronger than Zone 1 but not fully inland-type
- Hydration derived primarily from intermittent fog pulses, dew events, and short-term moisture retention in shallow soils
Examples: Transitional Copiapoa cinerea from Cifuncho, Esmeralda, upper Taltal, and mid-slope Paposo hinterland populations, along with transitional forms of humilis and fiedleriana.
Primary anchors: Taltal (coastal to transitional), Paposo hinterland, Caleta Cifuncho, Caleta Esmeralda, Pan de Azucar (transitional), Llanos de Challe (transitional).
➤ Zone 3: Inland Fog-Shadow Forms (700 to 1,100 m)
Environmental context: Fog may form episodically but contributes
little usable moisture. Fog-water yield declines sharply, humidity events are brief, and plants function primarily as inland xerophytes. UV exposure increases and thermal amplitude becomes pronounced. Occasional humidity pulses do not offset the long-term decline in reliable fog input beyond the inversion zone.
Climatic data: Available moisture drops dramatically, with absolute humidity often falling into the 1 to 6 g/kg range at inland sites. Potential water collection from fog is far lower than in coastal environments.
Traits:
- Thin or patchy epicuticular wax limited to the youngest growth
- Darker epidermis in green, slate, grey, or near-black tones due to UV-induced pigments and thickened cuticles
- Compact globose or tight columnar bodies with limited rib expansion
- Dense, protective spination, often darker in color
- Thickened epidermal walls consistent with chronic drought stress
Examples: Inland Copiapoa cinerea including Cerro Perales populations, inland Taltal and Esmeralda forms, atacamensis at lower inland elevations, angustiflora, upper-elevation coquimbana, and high-site tenuissima.
Primary anchors: Cerro Perales, inland Taltal corridors, inland Esmeralda slopes.
➤ Zone 4: High Montane Forms (Above 1,100 m)
Environmental context: This zone lies above the elevation where fog-water yield effectively collapses. Fog may be visible at times but contributes negligible hydration. Plants experience extreme UV exposure, very low humidity, strong winds, and wide diurnal temperature swings. Only a small fraction of Copiapoa individuals can establish and persist, resulting in sparse and highly localized populations under intense selection.
Climatic data: While the marine inversion layer blocks daytime fog, adiabatic cooling can drive nighttime relative humidity toward saturation on exposed peaks near the fog ceiling. This effect occurs within a narrow elevation band and may provide brief nocturnal moisture inputs despite the collapse of regular fog-water yield.
Lithic safety net: Possible contributions from mineral substrates that condense or retain trace moisture remain speculative and have not been experimentally demonstrated in Copiapoa.
Traits:
- Extremely compact, ultra-slow-growing bodies
- Deeply recessed apices protected by ribs and spines
- Bronze or olive coloration from UV-protective pigments visible through thin wax layers
- Thickened walls and reduced rib counts consistent with chronic drought stress
- Rigid, upright spines providing thermal and UV shielding
Examples: Rare montane populations of Copiapoa cinerea, angustiflora, and high-elevation coquimbana.
Primary anchor: Quebrada Botija, Cerro Perales (fog-edge and inversion-threshold reference).
Summary: From Zone 1 to Zone 4, Copiapoa shift from fog-dependent atmospheric hydration to extreme reliance on brief humidity pulses and mineral-bound moisture. The visible changes in form track this hydrological gradient more closely than any taxonomic boundary.
Note on overlaps and anchors: Numerical overlaps between zones represent ecotones where environmental conditions blend over short distances. Some anchors are included for diagnostic value in marking transitions or absences rather than continuous population corridors.
Source Basis: Zone boundaries integrate fog climatology from Cereceda et al. (2008) and Moat et al. (2021) with biological indicators documented across the coastal Atacama. Full citations are on the Reference page.
With the four core ecotypes defined, the next question is why two plants at the same elevation can look completely different. The answer lies in microhabitat. Elevation sets the ecotype, but substrate, slope, mineral chemistry, and terrain refine expression. These cross-elevation modifiers explain persistent variation within zones even when climate is shared.
Ecotype Zone Map
The interactive Copiapoa Ecotype Zone Map illustrates the coastal fog belt, mid-elevation transitions, fog-shadow interiors, high montane zones, and modifier pockets. This framework applies across the entire genus and explains why plants sharing a single name may behave very differently in cultivation.

Geologic mosaic underlying Copiapoa Atacama Desert habitat
Cross-Elevation Modifiers
Ecotype zones describe broad climatic environments: fog immersion, humidity stability, UV exposure, thermal regime, and water-use strategy. Within any ecotype zone, local microhabitat conditions can still produce striking differences in plant appearance.
Cross-elevation modifiers explain those differences. They do not redefine ecotype identity. Instead, they modify expression within an ecotype by changing heat load, surface reflectivity, rooting opportunity, nutrient availability, and boundary-layer microclimate.
A simple way to read the hierarchy is: Fog sets the water. Rock sets the heat. Microclimate shapes the plant. Surface constraints determine where it can root. Nutrient islands modify growth.
Geology, Albedo, and Substrate Memory
Fog corridors operate on top of an unusually persistent geological template. Minimal rainfall, low erosion rates, and long-term tectonic stability since at least the Miocene have preserved an exposed mineral mosaic across northern Chile.
In the hyper-arid Atacama Desert, this stability extends beyond the rock itself. Atmospheric inputs are not cycled out through rainfall or rapid biological turnover. Instead, salts, dust, atmospheric compounds, and biogenic inputs such as seabird guano can persist in place for thousands to millions of years (Ewing et al. 2006). Soil formation in these conditions is therefore less a process of renewal than one of accumulation. This creates a chemically stable but slowly evolving surface environment that reinforces the influence of underlying geology rather than obscuring it.
🔴 Startling observation: At most points along the desert coast, a traveler displaced a million years into the past would still recognize where they were.
Unlike most deserts, where soil development can homogenize substrates, the Atacama Desert presents abrupt transitions between granitic, volcanic, and mineralized surfaces over distances of only a few meters. These boundaries alter surface albedo, heat retention, and moisture dynamics. Because substrates remain stable over geological timescales, they act as long-term environmental switches, shaping Copiapoa morphology generation after generation within the same ecotype zone.
Short-term observation can be misleading. Fog presence varies by season and time of day, and surface appearance may conceal the true rooting substrate. A colony that appears to grow on pale, reflective sand may actually be rooted in dark volcanic rock only centimeters below the surface. These hidden substrate differences directly influence thermal load, moisture persistence, and long-term plant form.
Thermal Albedo and Microclimate
The thermal behavior of substrate is one of the main ways geology becomes biologically meaningful.
High-albedo granitic and alluvial surfaces reflect more incoming radiation. This can increase multidirectional light exposure around the plant while reducing heat retention in the substrate itself. Under these conditions, Copiapoa often express lighter bodies, stronger reflective wax, and straw to amber spine tones.
Low-albedo substrates, such as dark volcanic rock or iron-rich mineralized surfaces, absorb and re-radiate heat. This raises chronic thermal load around the stem and root zone, especially where nighttime heat release keeps the surface environment warm. Under these conditions, plants often express darker epidermis, stronger pigmentation, denser spination, and more compact growth.
This substrate-linked modulation of heat and radiation is the Thermal Albedo Effect. It explains how two plants in the same fog zone and elevation band can experience very different microclimates. One may grow in a cool, reflective, high-light environment; another may grow only meters away in a heat-retentive, low-albedo setting.
The key point is that substrate does not change the plant’s ecotype. It modifies how that ecotype is expressed.
Morphological Response: Spines, Wax, Ribs, and Pigment
Field and experimental studies show that Copiapoa orientation, rib geometry, surface reflectivity, and spine architecture function as active thermal adaptations rather than incidental growth traits.
Cactus spines are not simply defensive structures. Experimental and comparative studies show that they act as passive regulators of stem microclimate. Spine density, thickness, color, and orientation modify light interception, boundary-layer humidity, and surface temperature, directly influencing physiological performance under sustained stress.
Across Cactaceae, spine architecture responds strongly to long-term thermal and radiative environments. Spine-removal experiments consistently demonstrate increased stem temperatures and photoinhibition under high irradiance. Quantitative anatomical work also shows that spine traits can vary developmentally in response to environmental conditions rather than taxonomic identity alone.
In Copiapoa, where thermal and radiative stress vary sharply across fog belts and substrates, spine form, density, pigmentation, wax expression, and rib architecture are best interpreted as parts of an environmentally driven functional continuum rather than as simple indicators of genetic divergence.
Spine color also has limits. Environmental conditions influence the shade, density, and banding of newly formed spines through developmental responses to sustained thermal and radiative stress. Existing spines do not change color, and new spines vary only within the inherited range of the lineage. A dark-spined lineage will not produce pale yellow spines outside its inherited range without population-level evolutionary change or possible hybrid ancestry.
Where contrasting substrates meet, morphology may appear intermediate because microclimates blend across short distances. These gradients do not imply hybrid ancestry or taxonomic separation. They show how environmental modulation operates within genetic limits.
Surface Constraints and Rooting Environments
Surface substrates shape plant form indirectly by controlling where plants can establish and how roots access the substrate. The main substrate types in Copiapoa habitat are described below.
Caliche crusts
Caliche is a hardened subsurface crust formed by the accumulation of soluble salts such as calcium carbonate, gypsum, and sodium nitrate. In the Atacama Desert, caliche develops where evaporation exceeds precipitation and leaching is minimal. It is chemically restrictive and physically cemented, functioning more as a surface constraint than as usable soil.
Large caliche plains are well developed along the Chañaral–Caldera corridor, in interior basins near Llanos de Challe, and across ancient coastal terraces south of Pan de Azúcar. In these landscapes, Copiapoa usually persist along fracture lines, crust margins, and discontinuities where limited moisture and ions can accumulate.
Caliche does not supply water or nutrients in any simple beneficial sense. Its main effect is competitive exclusion. By suppressing most vegetation and limiting rooting depth, it creates open surfaces where stress-tolerant Copiapoa can persist. Dense colonies may form around the few fractures that allow access to underlying substrate.
In these settings, plant form remains governed by fog regime, thermal load, and bedrock properties. Caliche may shape rooting pattern and colony structure, but it does not override ecotype or substrate-driven microclimate.
Talus and scree slopes
Steep talus slopes, cliffs, and unstable scree intensify heat loading, reduce soil depth, and impose constant mechanical stress. Fog exposure depends strongly on slope aspect and airflow.
Plants in these conditions often develop leaning or creeping growth, anchoring into fractured rock rather than stable soil. They may show stronger outward or reflexed spines, thickened cuticles, reinforced ribs, and slower growth due to poor water retention.
Examples include talus-grown Copiapoa cinerea, cliff-bound C. solaris and C. serpentisulcata, and scree-based C. bridgesii. These forms are best understood as local expressions shaped by mechanical instability, heat load, and limited rooting opportunity within the broader ecotype framework.
Lithologic Contact Zones
Where contrasting substrates meet, plant expression can shift abruptly over very short distances.
Light granitic or alluvial surfaces tend to produce cooler, more reflective microclimates. Dark volcanic or iron-bearing substrates tend to produce hotter, more radiative microclimates. At contact zones, these conditions can change within a single slope or basin. Mineral chemistry may influence stress physiology and nutrient balance locally, but heat, UV exposure, and surface reflectivity remain the dominant drivers of visible phenotypic shifts.
Mineral presence should therefore be treated as contextual, not as a simple direct cause of color.
Localized Nutrient Modifiers
Most cross-elevation modifiers act through heat, reflectivity, rooting depth, or substrate stability. Biogenic nutrient islands operate differently. They do not change ecotype or define morphology directly; instead, they may locally modify growth conditions where nutrient sources, fog transport, and suitable rooting surfaces overlap.
Guano and biogenic nutrient islands
While most of the Atacama Desert is mineral-dominated and biologically sparse, certain coastal cliffs and headlands function as localized biogenic nutrient islands due to persistent seabird activity over late Pleistocene to Holocene timescales. In this hyper-arid environment, minimal precipitation, limited microbial turnover, and slow organic decomposition allow biogenic inputs to accumulate and persist, producing nitrogen- and phosphorus-enriched microsites distinct from surrounding mineral substrates.
The Humboldt Current sustains high marine productivity and large seabird colonies, whose long-term guano accumulation represents one of the few enduring biological nutrient subsidies in the coastal Atacama Desert. These inputs originate from marine trophic cycling and carry a distinct chemical and isotopic signature not present in terrestrial desert systems. Seabird guano from northern Chilean rookeries shows extremely elevated nitrogen isotope values (δ¹⁵N averaging ~23‰), producing levels of enrichment far beyond what aridity or natural soil processes can generate (Santana-Sagredo et al.). In fog-exposed Copiapoa habitats near former rookeries and historic guano extraction sites, roots may occur in proximity to guano-enriched debris and altered soil microbiomes, locally increasing nutrient availability.
Coastal fog (Camanchaca) interacting with these enriched surfaces can entrain dissolved nitrogenous compounds and aerosols. In Copiapoa, spine-mediated capture of atmospheric moisture is functionally integrated with internal water transport, allowing intercepted water to move from the plant surface into the stem.
Biogenic inputs may therefore contribute to nutrient availability both through localized deposition to the rhizosphere and through plant-surface interception. In this way, any nutrient contribution is incremental and occurs only where fog already provides reliable hydration. These effects are spatially constrained, typically limited to areas near active or historic seabird colonies, and diminish rapidly with distance. Guano enrichment therefore operates as a localized modifier rather than a landscape-scale driver of these systems.
The spatial structure of this process can be difficult to visualize from text alone. In coastal systems such as Guanillos, the interaction between fog transport, topography, and localized nutrient sources occurs over very short distances. The conceptual model below illustrates how these elements converge at the mouth of the quebrada.
Several historic guano localities illustrate this context. Guanillos (Huanillos) refers to more than one coastal site in northern Chile and must be carefully distinguished: the Tarapacá cliff system in the far north and the Antofagasta coastal cove near Cifuncho occur in different fog corridors and should not be conflated. Additional representative localities include Pabellón de Pica and Punta Gruesa. In some guano-influenced settings, nitrophilous lichens (e.g., Xanthoria spp.) occur on surrounding rock surfaces, providing a visible field indicator of localized nitrogen enrichment within otherwise mineral-dominated fog-belt habitats.
Biogenic nutrient islands should therefore be understood as contextual modifiers of growth potential within fog-supported ecotypes, not as primary drivers of Copiapoa distribution, ecotype formation, or taxonomic differentiation.
Geologic substrate mosaic across the Atacama Desert fog belt
This sample geologic map segment, based on the Mapa Geologico de Chile (SERNAGEOMIN 2003), shows the mineral patchwork underlying Copiapoa habitats. The map was developed for mineral and structural mapping but offers unusually high resolution of the environmental template underlying Copiapoa distribution. In much of the coastal Atacama Desert, ancient metamorphic basement, Jurassic volcanic units, and Quaternary alluvium remain fully exposed due to extreme aridity and minimal soil development. There is little or no organic topsoil to blur geological boundaries. What is visible at the surface is the substrate itself.
By aligning official map units such as Jag, Dc4, Qa, and granitic intrusions with the Thermal Albedo Effect, the combined influence of substrate reflectivity and heat retention on pigment expression becomes easier to interpret. Match map units to the substrate classes described below to anticipate thermal tendencies and likely phenotypic expression at a given locality. Read the geology, and you can often anticipate how plants from a site are likely to present in body color and spine tone before seeing them in habitat.

Topographic corridors and fog penetration
Where coastal escarpments are interrupted by quebradas or low-relief terrain, fog can penetrate inland more effectively. These breaks act as localized pathways for moisture movement, concentrating fog along drainage mouths and adjacent slopes.
In guano-influenced systems, this same pathway can transport dissolved nutrients and aerosols over short distances. The result is a tightly constrained zone where fog, topography, and nutrient input intersect, typically limited to coastal interfaces and diminishing rapidly inland.
Substrate Classes and Phenotypic Outcomes
The following substrate classes summarize how persistent surface conditions can shape expression within the four-zone framework.
➤ The Mirror Effect: Light Granitic and Alluvial Basins
Map units: Qa, CPg, TRg, Klag. Quaternary alluvium and quartz-rich granitoids present high-albedo surfaces that reflect radiation and remain relatively cool under solar exposure. Silica-rich, iron-poor substrates with minimal heat retention result in lower thermal stress, reducing selective pressure for dark pigmentation and allowing pale epidermis and lighter spines to persist. El Soldado and Taltal alluvial fans (KK 611, KP 821) and Llanos de Challe coastal plains consistently produce porcelain bodies and yellow spines. Golden columna-alba forms occur where granitic sands dominate.
➤ The Heat Battery: Dark Volcanic and Sedimentary Massifs
Map units: Jag, J3l, Dc4, MP1c. Jurassic volcanic and marine sediments and older metamorphic basement present low-albedo surfaces that absorb and re-radiate heat, elevating stem and root-zone temperatures. Higher iron and magnesium content increases heat storage and UV stress, with darker epidermis and heavier spination associated with chronic thermal and UV stress. The Cifuncho and Esmeralda volcanic belts reliably produce melanistic cinerea expressions. Jet-black populations are documented around RMSD 189 and KK 612.
➤ The Black Battery: Extreme Iron-Oxide Mineralization
Map indicators: Fe and Cu anomalies within volcanic units. Iron-rich surfaces create localized super-heated microclimates even as thin surface layers. High iron availability may amplify stress responses, though direct elemental mechanisms remain untested and should not be assumed as causal. The darkest expressions are observed here: jet black, bronze-black, and deep graphite pigmentation. Documented in the Manto Huanillo mineral belt (KP 833 to 836; KK 1396). Isolated interior sites such as Quebrada Botija show that extreme dark phenotypes can arise in fog-shadow refugia where mineral stress and UV load dominate despite limited fog input.
➤ The Flip Zone: Geological Contact Boundaries
Where light granitic substrates meet dark volcanic or iron-bearing rock, phenotype can shift abruptly over very short distances. Along the Tigrillo to Las Lozas corridor, yellow columna-alba forms transition sharply into darker pigmentation at the precise point where basement geology changes. Mineral chemistry may influence stress physiology and nutrient balance locally, but heat, UV exposure, and surface reflectivity remain the dominant drivers of phenotypic shifts. Mineral presence should be treated as a contextual factor, not a direct cause of color.
How Cross-Elevation Modifiers Fit the Four-Zone System
Cross-elevation modifiers shape expression, not identity. A Zone 1 plant on reflective granite may appear golden. A Zone 2 plant on hot scree may appear darker and compact. A Zone 3 plant on volcanic rock may appear nearly black. A Zone 4 plant in shaded ravines may remain greener than exposed neighbors. These differences arise from persistent microclimatic forcing, not species divergence.
Conceptual model of fog-mediated nutrient transport in a coastal alluvial fan system (Guanillos)
Geomorphic setting (alluvial fan geometry, coastal cliff, and quebrada structure) based on Walk et al. (2019). Seabird guano accumulation at coastal rookeries provides a localized nutrient source.
Transport of dissolved nitrogen compounds and fine particulates via fog (Camanchaca), and their deposition at the quebrada mouth and upper fan slopes, is presented as a proposed mechanism consistent with short transport distances (~1.2 km) and established fog dynamics, but has not been directly measured in this system.
This model illustrates how biogenic inputs can function as localized modifiers within fog-supported ecotypes, influencing growth conditions without altering the underlying ecological structure.
Why Copiapoa Cinerea Stands Alone
Copiapoa cinerea is among the most ecologically wide-ranging lineages in the genus. It spans persistent coastal fog belts, drier interior transitions, and higher slope environments. This unusually broad habitat amplitude explains several otherwise puzzling features of the complex: extraordinary phenotypic diversity, the historical proliferation of locality and form names including columna-alba and krainziana, consistently low genetic divergence reported in molecular studies, and a comparatively more stable conservation outlook relative to narrow endemics such as Copiapoa laui, esmeraldana, and tenuissima.
The diversity within Copiapoa cinerea is best explained as long-term ecological differentiation within recurring environmental corridors rather than cryptic speciation or widespread hybridization. The extreme forms collectors prize represent ecotype expression, not separate species. Only plants originating from the correct ecotype zone can express the full expected range of morphology associated with that ecotype. Cultivation alone cannot convert one ecotype into another.
Molecular phylogenetic sampling shows that Copiapoa cinerea and its columna-alba and krainziana forms constitute a single genetic lineage with low divergence, supporting their treatment as geographically structured ecotypic expressions rather than distinct species (Larridon et al. 2015). Nuclear microsatellite analyses in Larridon et al. 2018 corroborate this interpretation, with AMOVA results indicating that over 90% of detected genetic variation is distributed within named taxa rather than between them. The 2025 Sarnes monograph reinforces this interpretation through extensive habitat photography, documenting repeatable morphological expressions wherever environmental conditions align. Locality-based ecology provides the most coherent explanation for cinerea diversity and for why plants bearing the same name may require very different cultivation regimes.
Why two plants labeled Copiapoa cinerea may differ
Two plants both sold as Copiapoa cinerea can look like different species. One may be porcelain white with soft ribs and short pale spines. The other may be near-black with dense dark spination and a compact, hardened body. Neither is mislabeled. The difference comes from three variables acting together.
First, ecotype zone. A coastal Zone 1 plant from the Taltal fog belt develops under persistent humidity, producing heavy pruina, broad hydrated ribs, and soft epidermis. An inland Zone 3 plant from Cerro Perales develops under chronic drought and high UV, producing darker pigmentation, compact growth, and thickened cuticle. These are different ecological strategies within the same lineage.
Second, substrate class. Within the same zone, a plant rooted over light granitic alluvium (Mirror Effect) will tend lighter in body and spine color than one rooted over dark volcanic rock (Heat Battery) only meters away. Substrate controls thermal load and reflectivity at the root zone and stem surface, shifting expression without changing ecotype.
Third, provenance. A plant grown from habitat seed collected at a documented locality carries the genetic and developmental history of that population. A plant grown from cultivated seed of uncertain parentage may express traits from multiple populations or even undocumented hybrid crosses. Provenance determines whether the plant's expression can be meaningfully interpreted at all.
When all three variables are known, the differences between two cinerea plants become predictable rather than confusing. When any of the three is missing, confident interpretation is not possible.
From framework to practice: collector guidance
The diversity seen in Copiapoa is best understood as sustained ecological differentiation along persistent environmental corridors rather than as rigid species boundaries. To achieve habitat-correct results in cultivation, growers should work through three linked decisions.
➤ Atmospheric alignment: ecotype zone
Begin by placing the plant within its correct ecotype zone. High-humidity coastal belts in Zones 1 and 2 favor more hydrated, globose forms. Inland and high-montane environments in Zones 3 and 4 select for compact bodies, leathery epidermis, and deeply recessed apices under high UV and thermal stress. No amount of cultivation can make a Zone 3 inland clone express the long-term morphology of a Zone 1 coastal clone, or vice versa. Ecotype sets the physiological baseline.
➤ Thermal alignment: Thermal Albedo Effect
While fog regime defines the overall body plan, substrate controls thermal load and therefore pigmentation and spine expression. High-albedo mirror substrates such as light granitic sands reflect solar radiation and favor silver-white bodies with straw to amber spines.
Low-albedo heat-battery and black-battery substrates such as dark volcanic rock and iron-rich mineralization absorb and re-radiate heat, promoting darker epidermis and denser, darker spination typical of inland melanohystrix-type expressions.
In cultivation, this explains why plants grown on pale, cool mineral mixes tend to remain lighter, while those on darker, heat-retentive media intensify pigmentation under the same light regime.
➤ Functional biology: living mineral matrix
Atmospheric and thermal alignment only succeed if the root zone can support the imposed stress regime. In habitat, inland and high-montane ecotypes do not grow in sterile soil but within a living mineral matrix. Metagenomic sampling of Copiapoa solaris rhizosphere communities across the Quebrada Botija corridor documents that microbial community structure and stress-response gene abundance differ meaningfully across populations separated by as little as 25 km, tracking local humidity gradients and thermal load (Cayo et al. 2025).
These findings support the interpretation that Copiapoa ecotypes are embedded within long-term soil biological legacies that are themselves structured by the same environmental gradients shaping plant morphology. Successful cultivation of inland and high-elevation forms therefore likely depends on recreating not only mineral structure but also a functional microbial matrix capable of mediating nutrient exchange, osmotic stress, and micro-scale water access. Without this biological support, attempts to hard-grow Zone 3 and Zone 4 plants commonly result in slow decline rather than habitat-correct hardening.
🔴 Principle: Match ecotype zone to body plan, then align thermal substrate and functional biology. Only when all three are in place will Copiapoa reliably express the long-term, habitat-correct appearance associated with specific localities.
Copiapoa cinerea distribution and reference localities
This map presents a schematic overview of the Copiapoa cinerea lineage complex across the Atacama Desert coastal range, emphasizing core coastal corridors, recurring morphological expressions, and key reference localities. Polygon boundaries are schematic visualization bands and do not represent surveyed range limits.
Copiapoa gigantea occupies the northern corridor from roughly the Quebrada Izcuña area south toward Paposo, where coastal rocky slopes support the lineage's most massive columnar expressions. It is shown here as a related expression within the broader cinerea framework, noting its comparatively more distinct clustering signal in nuclear analyses (Larridon et al. 2015).
Copiapoa cinerea sensu stricto is centered on the Taltal coastal massif, the type corridor for the complex. High-elevation fog-belt ridges in this corridor define the baseline morphological benchmark against which other expressions are interpreted.
Copiapoa cinerea, krainziana form is shown as a geographically restricted ecotypic expression northeast of Taltal, centered on the San Ramón Valley hillside niche. It is treated here as a stable ecotypic expression within the cinerea lineage rather than a distinct species, consistent with the absence of sequence variation between these forms in plastid marker analyses.
Copiapoa cinerea, columna-alba form occupies the southernmost corridor of the complex, spanning the granite coastal shelf from south of Taltal toward Chañaral. The extreme pruina development associated with this expression reflects substrate and insolation conditions specific to this sector and is interpreted as an ecotypic response rather than a discrete genetic discontinuity.
Together these four expressions illustrate why two plants labeled Copiapoa cinerea may differ substantially in morphology and cultivation requirements. Locality and habitat context remain the most reliable guide to interpreting these plants correctly.
Source Basis: Low genetic divergence within the cinerea complex follows Larridon et al. (2015, 2018). Habitat-based morphological interpretation follows Sarnes (2025). Rhizosphere community structuring follows Cayo et al. (2025). Full citations are on the Reference page.
Practical Interpretation for Collectors and Conservation
The framework presented in this guide, ecotype zones, cross-elevation modifiers, geographic anchors, and the cinerea case study, converges on a small number of practical principles. These apply whether you are evaluating a plant for purchase, labeling material for sale, interpreting habitat photography, or assessing conservation priorities.
Provenance first
No amount of morphological analysis can substitute for reliable provenance. A plant with documented locality, collector number, and seed history can be interpreted within the ecotype framework. A plant without that information cannot, regardless of how closely it resembles a known form. When provenance is missing, say so. "Unknown origin" is more useful than a confident but unsupported name.
Do not confuse ecotype variation with hybrid ancestry
Plants from different ecotype zones within the same lineage can look radically different. A coastal Zone 1 Copiapoa cinerea and an inland Zone 3 Copiapoa cinerea may share a name but differ in body color, wax, spination, rib structure, and growth rate. These differences reflect long-term environmental forcing, not hybridization. Before suspecting hybrid origin, check whether the plant's traits are consistent with its documented ecotype zone and substrate class. Most variation that looks "intermediate" or "unusual" has an ecological explanation when habitat context is available.
Do not confuse habitat plants with habitat seed
A wild-collected plant growing in cultivation is a habitat plant. Seed produced by that plant in cultivation is not habitat seed unless pollination history is documented. Open pollination in mixed collections is one of the most common sources of cryptic hybridization in Copiapoa. Seed provenance requires the same level of documentation as plant provenance.
Label uncertainty honestly
When origin is unclear, traits appear inconsistent, or documentation is incomplete, label the plant conservatively. "Suspected hybrid," "uncertain origin," or "cultivated seed, parent locality X" are all more responsible than assigning a species name with false confidence. Honest labeling protects the integrity of cultivated seed pools and preserves the value of well-documented material.
Use anchors, zones, and substrate together
No single variable explains Copiapoa morphology on its own. Ecotype zone sets the baseline architecture and water-use strategy. Substrate class modifies expression through thermal load and reflectivity. Geographic anchor provides the landscape context that connects a locality to its fog corridor, elevation band, and geological setting. Interpretation is strongest when all three are used together and weakest when any one is treated as sufficient on its own.
Closing principle
Hybridization may blur boundaries in nature, but records must remain precise. Ecotype variation may blur appearances, but habitat context restores meaning.
The interactive Copiapoa Distribution Map below applies this framework across the full genus range, showing how ecotype zones, geographic anchors, and substrate corridors organize the diversity described throughout this guide.

The amazing diversity of Copiapoa cinerea